

Dynamin recruitment and membrane scission at the neck of a clathrin-coated pit
- PMID: 25232009
- PMCID: PMC4230619
- DOI: 10.1091/mbc.E14-07-1240
Dynamin recruitment and membrane scission at the neck of a clathrin-coated pit
Abstract
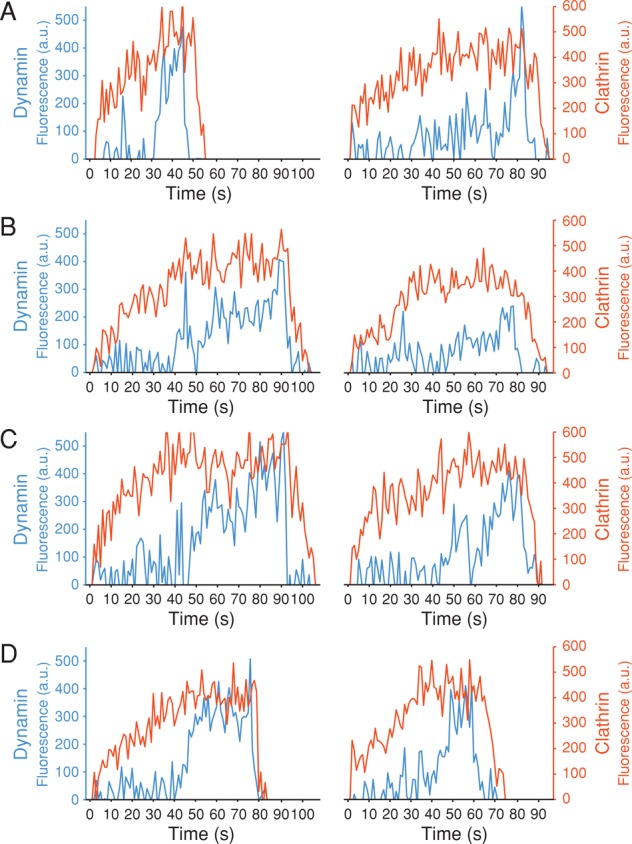
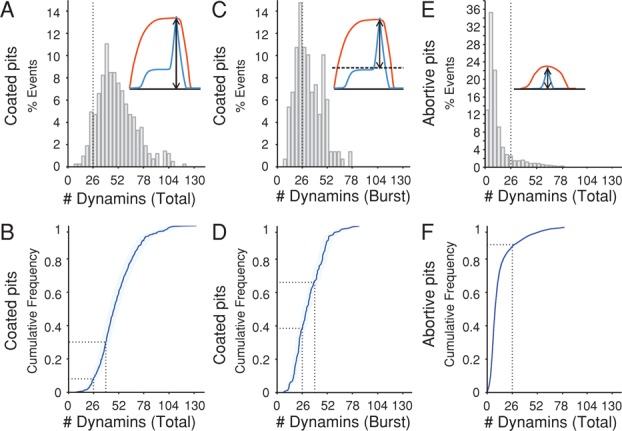
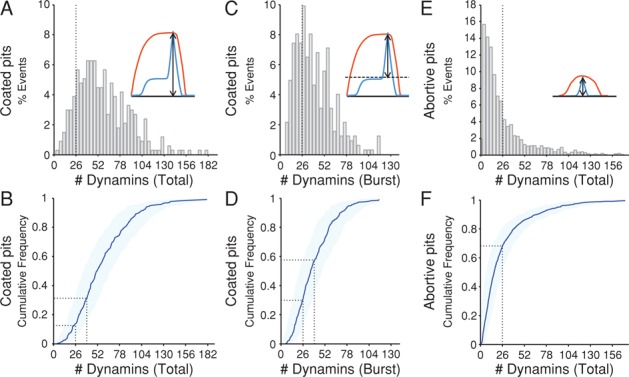
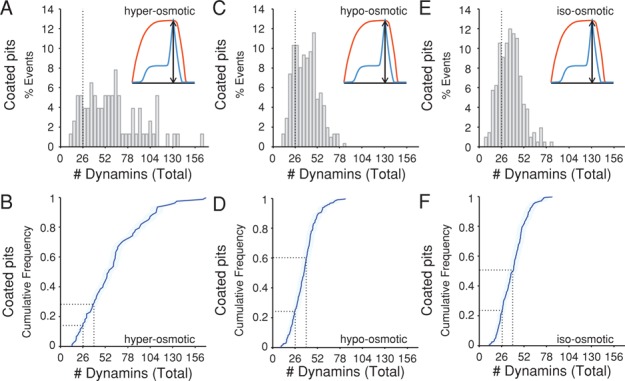
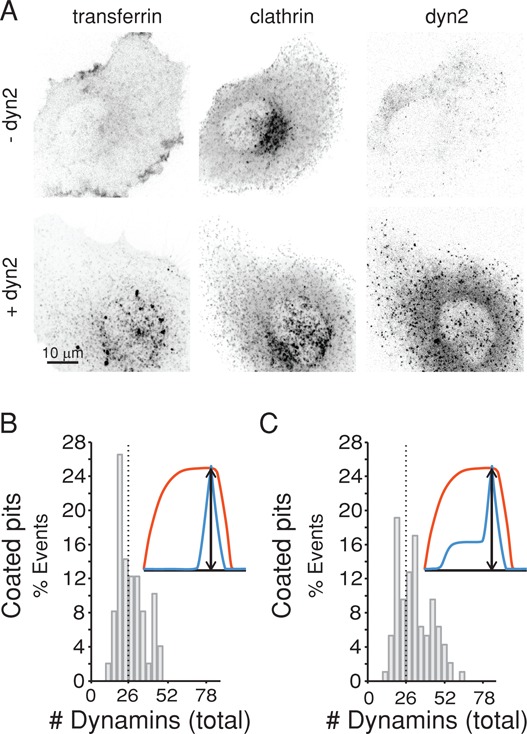
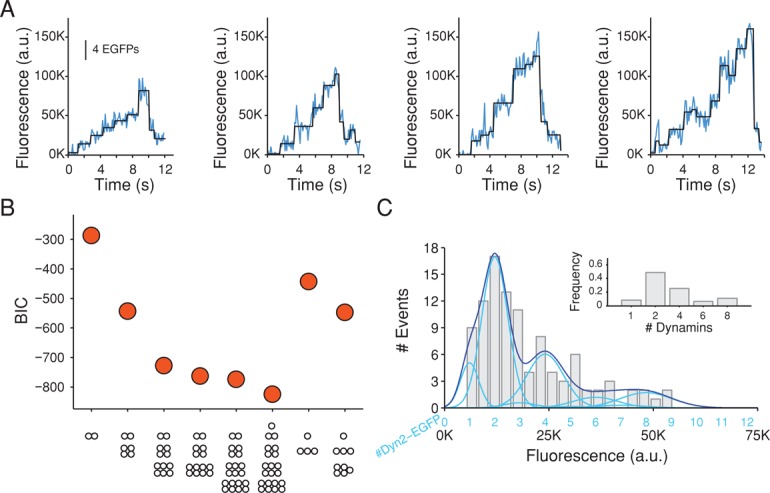
Dynamin, the GTPase required for clathrin-mediated endocytosis, is recruited to clathrin-coated pits in two sequential phases. The first is associated with coated pit maturation; the second, with fission of the membrane neck of a coated pit. Using gene-edited cells that express dynamin2-EGFP instead of dynamin2 and live-cell TIRF imaging with single-molecule EGFP sensitivity and high temporal resolution, we detected the arrival of dynamin at coated pits and defined dynamin dimers as the preferred assembly unit. We also used live-cell spinning-disk confocal microscopy calibrated by single-molecule EGFP detection to determine the number of dynamins recruited to the coated pits. A large fraction of budding coated pits recruit between 26 and 40 dynamins (between 1 and 1.5 helical turns of a dynamin collar) during the recruitment phase associated with neck fission; 26 are enough for coated vesicle release in cells partially depleted of dynamin by RNA interference. We discuss how these results restrict models for the mechanism of dynamin-mediated membrane scission.
© 2014 Cocucci et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
Similar articles
-
A noncanonical role for dynamin-1 in regulating early stages of clathrin-mediated endocytosis in non-neuronal cells.PLoS Biol. 2018 Apr 18;16(4):e2005377. doi: 10.1371/journal.pbio.2005377. eCollection 2018 Apr. PLoS Biol. 2018. PMID: 29668686 Free PMC article.
-
The role of dynamin and its binding partners in coated pit invagination and scission.J Cell Biol. 2001 Jan 22;152(2):309-23. doi: 10.1083/jcb.152.2.309. J Cell Biol. 2001. PMID: 11266448 Free PMC article.
-
Divergent modes for cargo-mediated control of clathrin-coated pit dynamics.Mol Biol Cell. 2013 Jun;24(11):1725-34, S1-12. doi: 10.1091/mbc.E12-07-0550. Epub 2013 Mar 27. Mol Biol Cell. 2013. PMID: 23536704 Free PMC article.
-
Building a fission machine--structural insights into dynamin assembly and activation.J Cell Sci. 2013 Jul 1;126(Pt 13):2773-84. doi: 10.1242/jcs.108845. Epub 2013 Jun 18. J Cell Sci. 2013. PMID: 23781021 Free PMC article. Review.
-
"Wunder" F-BAR domains: going from pits to vesicles.Cell. 2007 May 18;129(4):655-7. doi: 10.1016/j.cell.2007.05.006. Cell. 2007. PMID: 17512400 Review.
Cited by
-
How sticky? How tight? How hot? Imaging probes for fluid viscosity, membrane tension and temperature measurements at the cellular level.Int J Biochem Cell Biol. 2022 Dec;153:106329. doi: 10.1016/j.biocel.2022.106329. Epub 2022 Nov 3. Int J Biochem Cell Biol. 2022. PMID: 36336304 Free PMC article. Review.
-
Direct comparison of clathrin-mediated endocytosis in budding and fission yeast reveals conserved and evolvable features.Elife. 2019 Dec 12;8:e50749. doi: 10.7554/eLife.50749. Elife. 2019. PMID: 31829937 Free PMC article.
-
Total Internal Reflection Fluorescence (TIRF) Microscopy.Curr Protoc. 2022 Aug;2(8):e517. doi: 10.1002/cpz1.517. Curr Protoc. 2022. PMID: 35972209 Free PMC article. Review.
-
Dynamin 1xA interacts with Endophilin A1 via its spliced long C-terminus for ultrafast endocytosis.EMBO J. 2024 Aug;43(16):3327-3357. doi: 10.1038/s44318-024-00145-x. Epub 2024 Jun 21. EMBO J. 2024. PMID: 38907032 Free PMC article.
-
Regulation of dynamin family proteins by post-translational modifications.J Biosci. 2017 Jun;42(2):333-344. doi: 10.1007/s12038-017-9680-y. J Biosci. 2017. PMID: 28569256 Review.
References
-
- Binns DD, Barylko B, Grichine N, Atkinson MA, Helms MK, Jameson DM, Eccleston JF, Albanesi JP. Correlation between self-association modes and GTPase activation of dynamin. J Protein Chem. 1999;18:277–290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
