

Intracellular accumulation of amyloid-β (Aβ) protein plays a major role in Aβ-induced alterations of glutamatergic synaptic transmission and plasticity
- PMID: 25232124
- PMCID: PMC6705320
- DOI: 10.1523/JNEUROSCI.1201-14.2014
Intracellular accumulation of amyloid-β (Aβ) protein plays a major role in Aβ-induced alterations of glutamatergic synaptic transmission and plasticity
Abstract
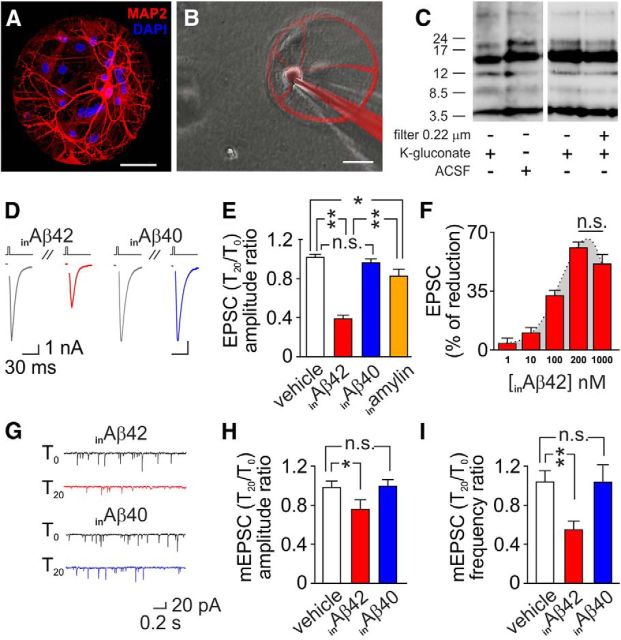
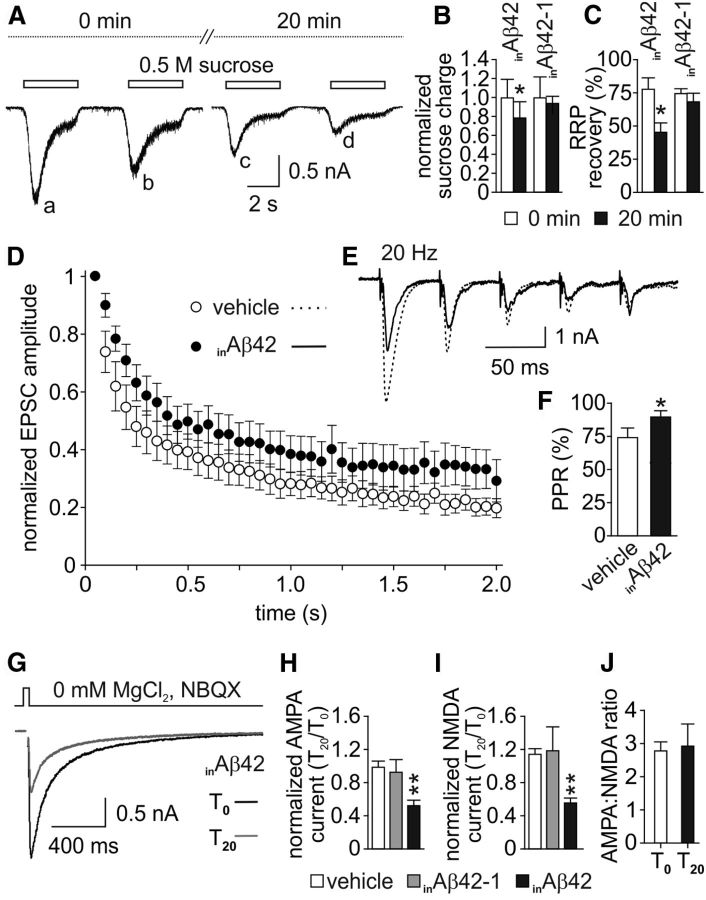
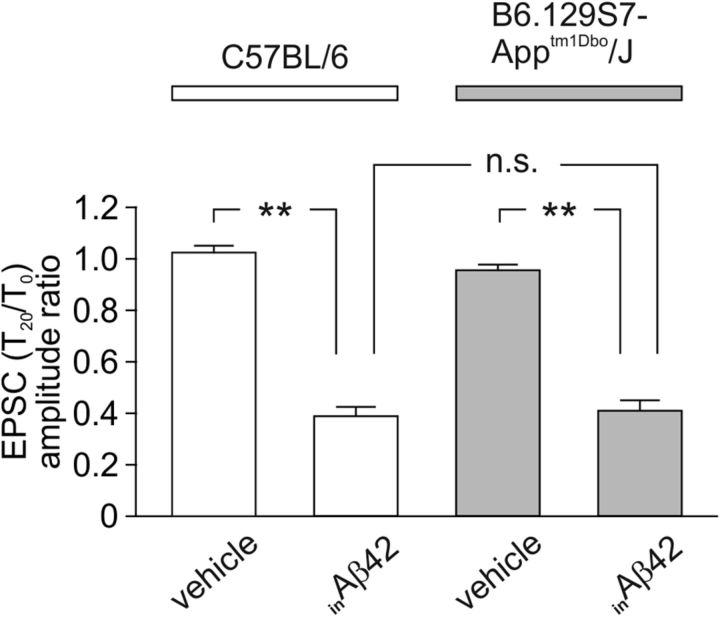
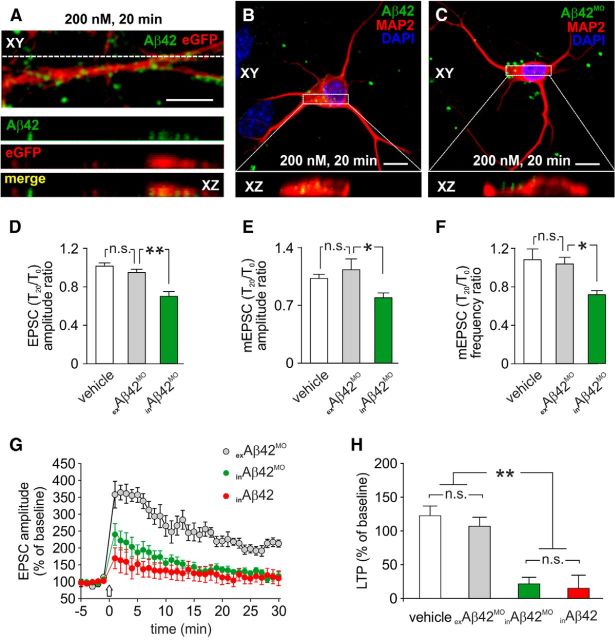
Intracellular accumulation of amyloid-β (Aβ) protein has been proposed as an early event in AD pathogenesis. In patients with mild cognitive impairment, intraneuronal Aβ immunoreactivity was found especially in brain regions critically involved in the cognitive deficits of AD. Although a large body of evidence demonstrates that Aβ42 accumulates intraneuronally ((in)Aβ), the action and the role of Aβ42 buildup on synaptic function have been poorly investigated. Here, we demonstrate that basal synaptic transmission and LTP were markedly depressed following Aβ42 injection into the neuron through the patch pipette. Control experiments performed with the reverse peptide (Aβ42-1) allowed us to exclude that the effects of (in)Aβ depended on changes in oncotic pressure. To further investigate (in)Aβ synaptotoxicity we used an Aβ variant harboring oxidized methionine in position 35 that does not cross the neuronal plasma membrane and is not uploaded from the extracellular space. This Aβ42 variant had no effects on synaptic transmission and plasticity when applied extracellularly, but induced synaptic depression and LTP inhibition after patch-pipette dialysis. Finally, the injection of an antibody raised against human Aβ42 (6E10) in CA1 pyramidal neurons of mouse hippocampal brain slices and autaptic microcultures did not, per se, significantly affect LTP and basal synaptic transmission, but it protected against the toxic effects of extracellular Aβ42. Collectively, these findings suggest that Aβ42-induced impairment of glutamatergic synaptic function depends on its internalization and intracellular accumulation thus paving the way to a systemic proteomic analysis of intracellular targets/partners of Aβ42.
Keywords: 6E10; amyloid-β protein; autaptic hippocampal neurons; intraneuronal accumulation; synaptic transmission; whole-cell LTP.
Copyright © 2014 the authors 0270-6474/14/3412893-11$15.00/0.
Figures



References
-
- Attar A, Ripoli C, Riccardi E, Maiti P, Li Puma DD, Liu T, Hayes J, Jones MR, Lichti-Kaiser K, Yang F, Gale GD, Tseng CH, Tan M, Xie CW, Straudinger JL, Klärner FG, Schrader T, Frautschy SA, Grassi C, Bitan G. Protection of primary neurons and mouse brain from Alzheimer's pathology by molecular tweezers. Brain. 2012;135:3735–3748. doi: 10.1093/brain/aws289. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous