

Adaptation of distortion product otoacoustic emissions predicts susceptibility to acoustic over-exposure in alert rabbits
- PMID: 25234992
- PMCID: PMC4167750
- DOI: 10.1121/1.4868389
Adaptation of distortion product otoacoustic emissions predicts susceptibility to acoustic over-exposure in alert rabbits
Abstract
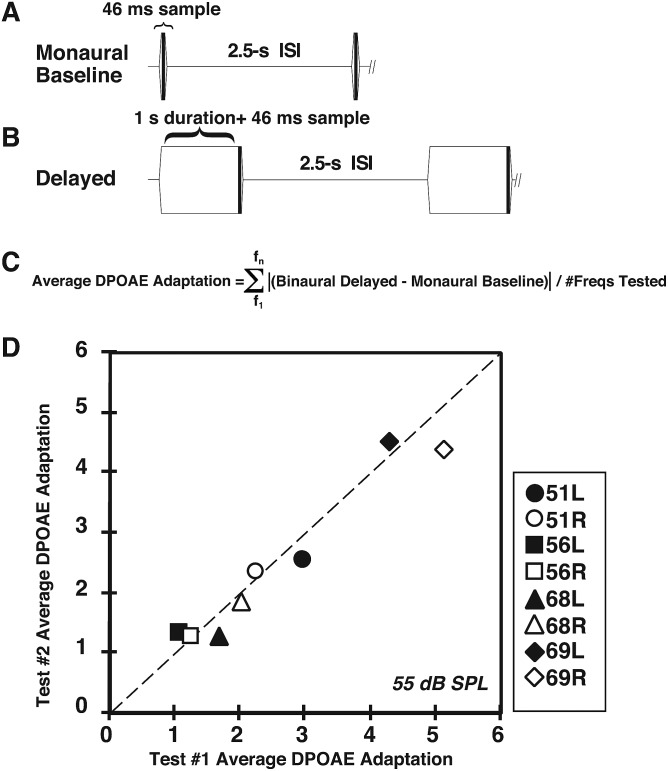
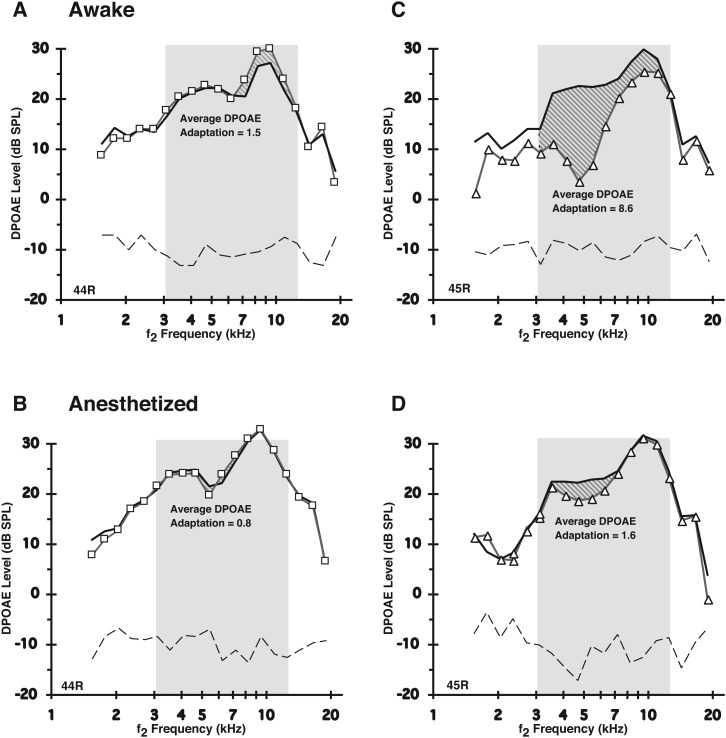
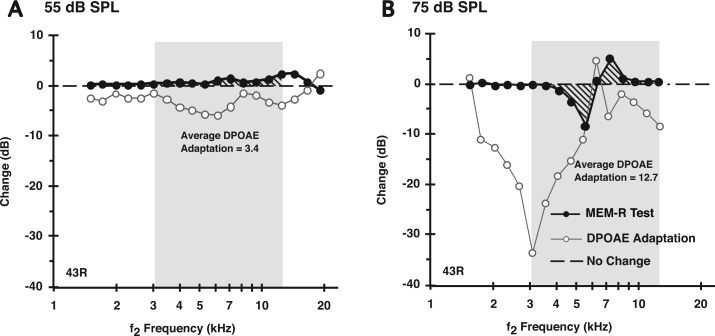
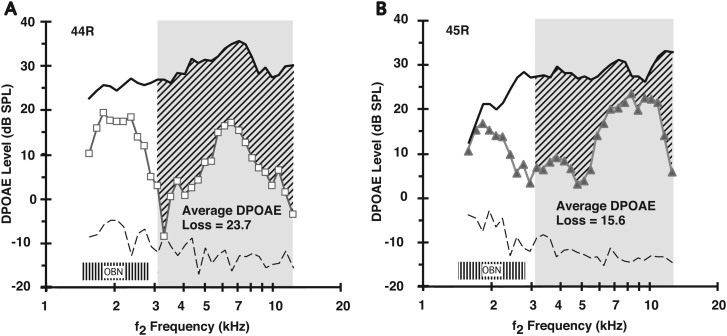
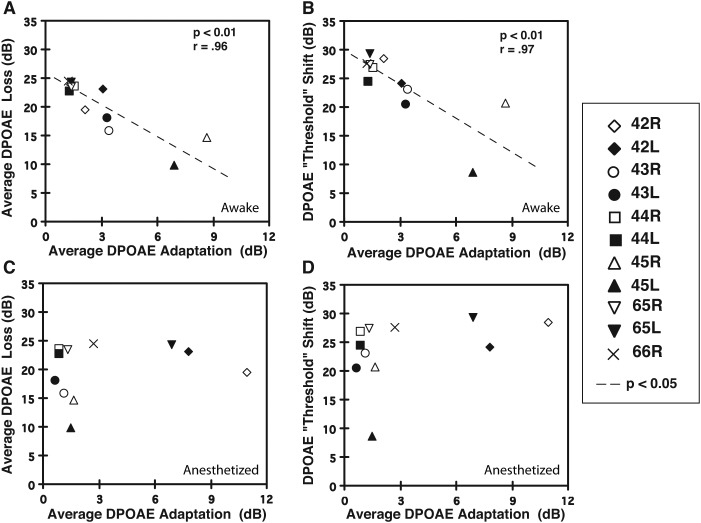
A noninvasive test was developed in rabbits based on fast adaptation measures for 2f1-f2 distortion-product otoacoustic emissions (DPOAEs). The goal was to evaluate the effective reflex activation, i.e., "functional strength," of both the descending medial olivocochlear efferent reflex (MOC-R) and the middle-ear muscle reflex (MEM-R) through sound activation. Classically, it is assumed that both reflexes contribute toward protecting the inner ear from cochlear damage caused by noise exposure. The DP-gram method described here evaluated the MOC-R effect on DPOAE levels over a two-octave (oct) frequency range. To estimate the related activation of the middle-ear muscles (MEMs), the MEM-R was measured by monitoring the level of the f1-primary tone throughout its duration. Following baseline measures, rabbits were subjected to noise over-exposure. A main finding was that the measured adaptive activity was highly variable between rabbits but less so between the ears of the same animal. Also, together, the MOC-R and MEM-R tests showed that, on average, DPOAE adaptation consisted of a combined contribution from both systems. Despite this shared involvement, the amount of DPOAE adaptation measured for a particular animal's ear predicted that ear's subsequent susceptibility to the noise over-exposure for alert but not for deeply anesthetized rabbits.
Figures
Similar articles
-
Influence of sound-conditioning on noise-induced susceptibility of distortion-product otoacoustic emissions.J Acoust Soc Am. 2015 Jul;138(1):58-64. doi: 10.1121/1.4922223. J Acoust Soc Am. 2015. PMID: 26233006 Free PMC article.
-
A multifrequency method for determining cochlear efferent activity.J Assoc Res Otolaryngol. 2002 Mar;3(1):16-25. doi: 10.1007/s101620010089. J Assoc Res Otolaryngol. 2002. PMID: 12083721 Free PMC article.
-
Aging of the medial olivocochlear reflex and associations with speech perception.J Acoust Soc Am. 2014 Feb;135(2):754-65. doi: 10.1121/1.4861841. J Acoust Soc Am. 2014. PMID: 25234884 Free PMC article.
-
Olivocochlear efferents: anatomy, physiology, function, and the measurement of efferent effects in humans.Ear Hear. 2006 Dec;27(6):589-607. doi: 10.1097/01.aud.0000240507.83072.e7. Ear Hear. 2006. PMID: 17086072 Review.
-
Morphofunctional alterations in the olivocochlear efferent system of the genetic audiogenic seizure-prone hamster GASH:Sal.Epilepsy Behav. 2017 Jun;71(Pt B):193-206. doi: 10.1016/j.yebeh.2016.05.040. Epub 2016 Aug 1. Epilepsy Behav. 2017. PMID: 27492627 Review.
Cited by
-
Otoacoustic-emission-based medial-olivocochlear reflex assays for humans.J Acoust Soc Am. 2014 Nov;136(5):2697-713. doi: 10.1121/1.4896745. J Acoust Soc Am. 2014. PMID: 25373970 Free PMC article.
-
Reflex Modification Audiometry Reveals Dual Roles for Olivocochlear Neurotransmission.Front Cell Neurosci. 2017 Nov 22;11:361. doi: 10.3389/fncel.2017.00361. eCollection 2017. Front Cell Neurosci. 2017. PMID: 29213229 Free PMC article.
-
Cochlear hair cell densities in the rabbit.Anat Sci Int. 2019 Jan;94(1):144-149. doi: 10.1007/s12565-018-0461-y. Epub 2018 Oct 1. Anat Sci Int. 2019. PMID: 30276665 Free PMC article.
-
Long term changes to auditory sensitivity following blast trauma in mice.Hear Res. 2021 Apr;403:108201. doi: 10.1016/j.heares.2021.108201. Epub 2021 Feb 12. Hear Res. 2021. PMID: 33636682 Free PMC article.
-
Influence of sound-conditioning on noise-induced susceptibility of distortion-product otoacoustic emissions.J Acoust Soc Am. 2015 Jul;138(1):58-64. doi: 10.1121/1.4922223. J Acoust Soc Am. 2015. PMID: 26233006 Free PMC article.
References
-
- Guinan, J. J. , Jr., Backus, B. C. , Lilaonitkul, W., and Aharonson, V. (2003). “Medial olivocochlear efferent reflex in humans: Otoacoustic emission (OAE) measurement issues and the advantages of stimulus frequency OAEs,” J. Assoc. Res. Otolaryngol. 4, 521–54010.1007/s10162-002-3037-3 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous