

Enzyme regulation. IRBIT is a novel regulator of ribonucleotide reductase in higher eukaryotes
- PMID: 25237103
- PMCID: PMC6428578
- DOI: 10.1126/science.1251550
Enzyme regulation. IRBIT is a novel regulator of ribonucleotide reductase in higher eukaryotes
Abstract
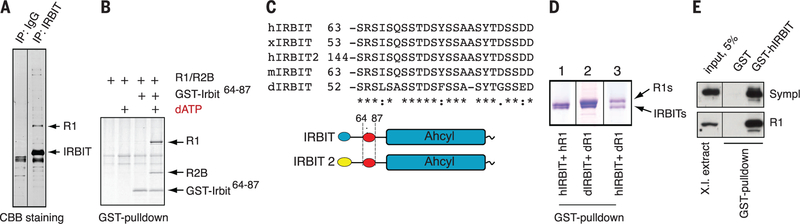
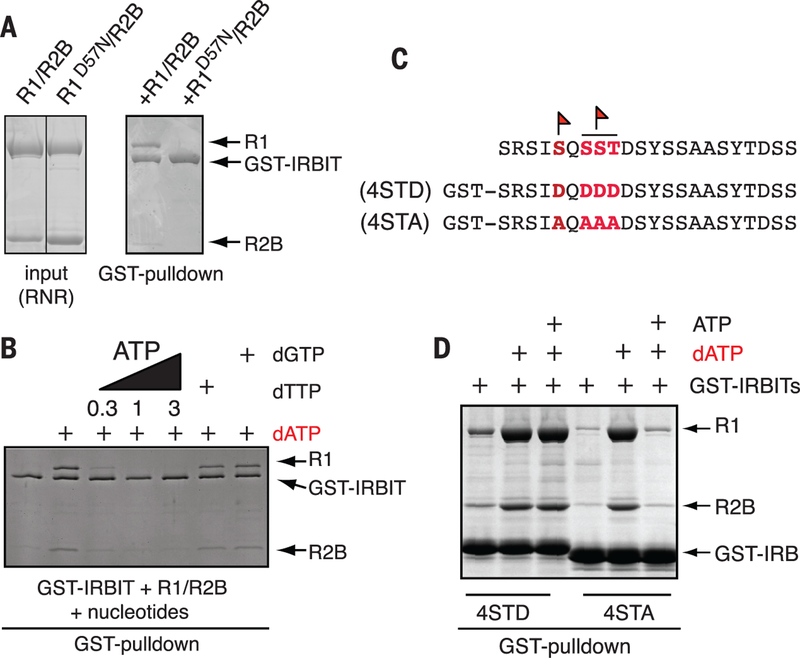
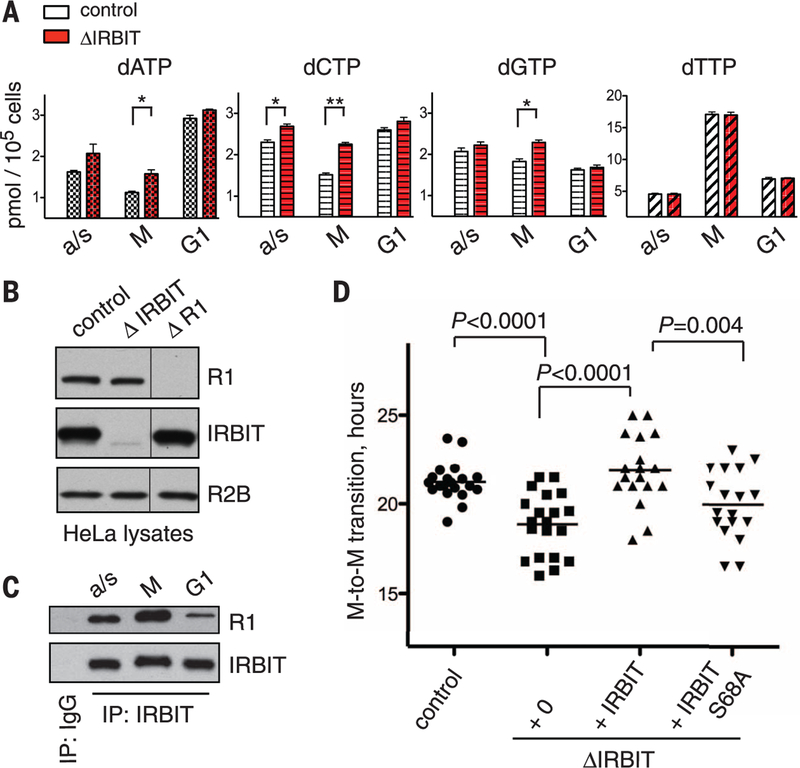
Ribonucleotide reductase (RNR) supplies the balanced pools of deoxynucleotide triphosphates (dNTPs) necessary for DNA replication and maintenance of genomic integrity. RNR is subject to allosteric regulatory mechanisms in all eukaryotes, as well as to control by small protein inhibitors Sml1p and Spd1p in budding and fission yeast, respectively. Here, we show that the metazoan protein IRBIT forms a deoxyadenosine triphosphate (dATP)-dependent complex with RNR, which stabilizes dATP in the activity site of RNR and thus inhibits the enzyme. Formation of the RNR-IRBIT complex is regulated through phosphorylation of IRBIT, and ablation of IRBIT expression in HeLa cells causes imbalanced dNTP pools and altered cell cycle progression. We demonstrate a mechanism for RNR regulation in higher eukaryotes that acts by enhancing allosteric RNR inhibition by dATP.
Copyright © 2014, American Association for the Advancement of Science.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
