

Cocaine inhibition of nicotinic acetylcholine receptors influences dopamine release
- PMID: 25237305
- PMCID: PMC4154440
- DOI: 10.3389/fnsyn.2014.00019
Cocaine inhibition of nicotinic acetylcholine receptors influences dopamine release
Abstract
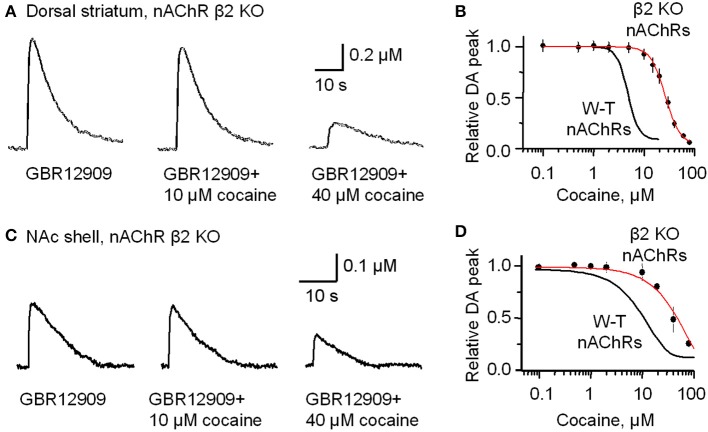
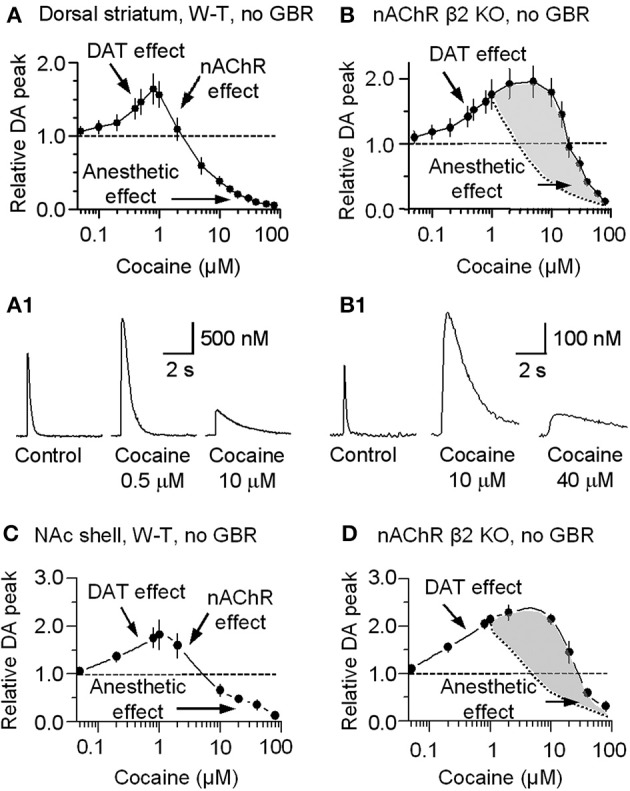
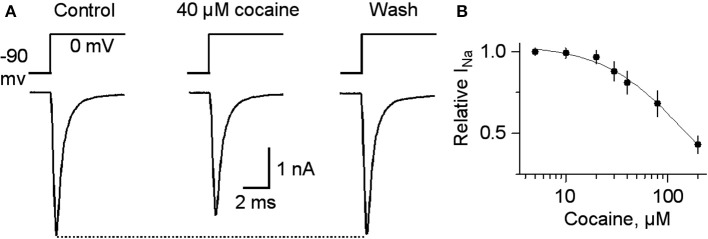
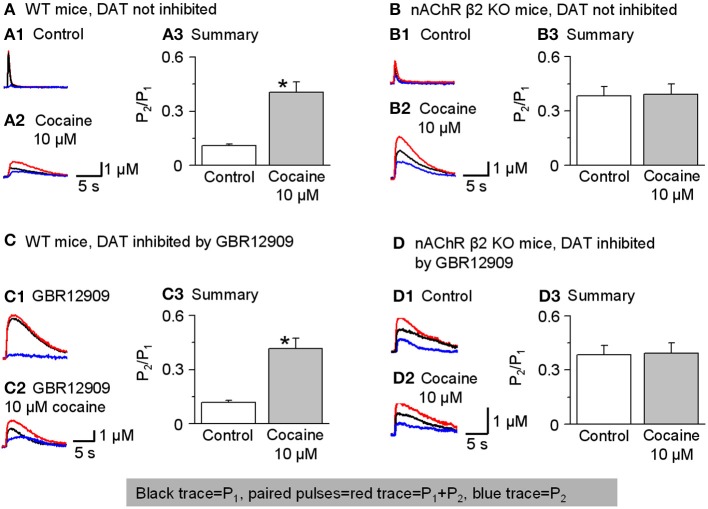
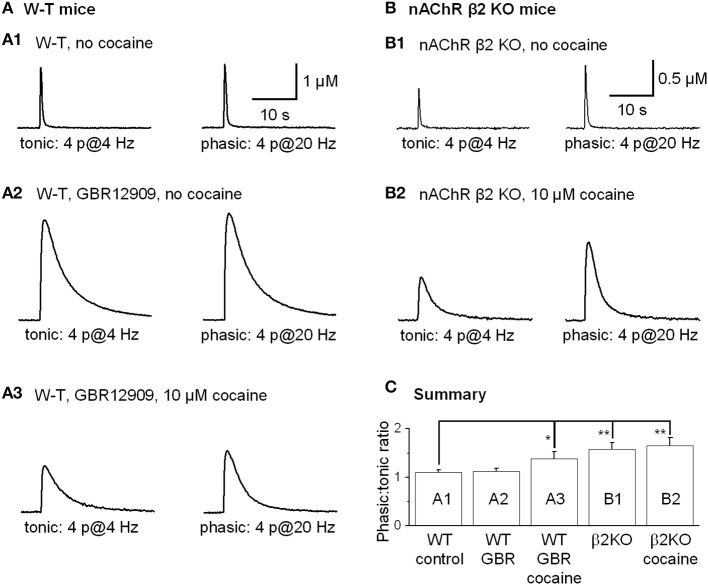
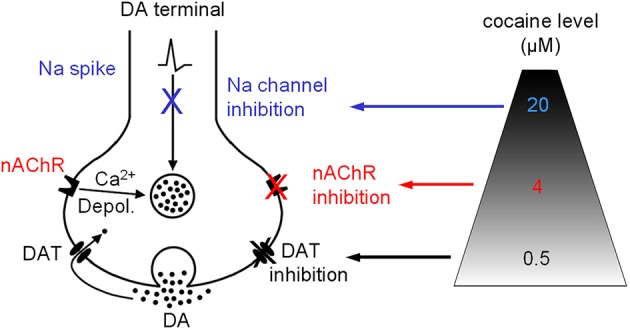
Nicotinic acetylcholine receptors (nAChRs) potently regulate dopamine (DA) release in the striatum and alter cocaine's ability to reinforce behaviors. Since cocaine is a weak nAChR inhibitor, we hypothesized that cocaine may alter DA release by inhibiting the nAChRs in DA terminals in the striatum and thus contribute to cocaine's reinforcing properties primarily associated with the inhibition of DA transporters. We found that biologically relevant concentrations of cocaine can mildly inhibit nAChR-mediated currents in midbrain DA neurons and consequently alter DA release in the dorsal and ventral striatum. At very high concentrations, cocaine also inhibits voltage-gated Na channels in DA neurons. Furthermore, our results show that partial inhibition of nAChRs by cocaine reduces evoked DA release. This diminution of DA release via nAChR inhibition more strongly influences release evoked at low or tonic stimulation frequencies than at higher (phasic) stimulation frequencies, particularly in the dorsolateral striatum. This cocaine-induced shift favoring phasic DA release may contribute to the enhanced saliency and motivational value of cocaine-associated memories and behaviors.
Keywords: addiction; mesolimbic; nAChRs; substantia nigra; ventral tegmental area; voltammetry.
Figures
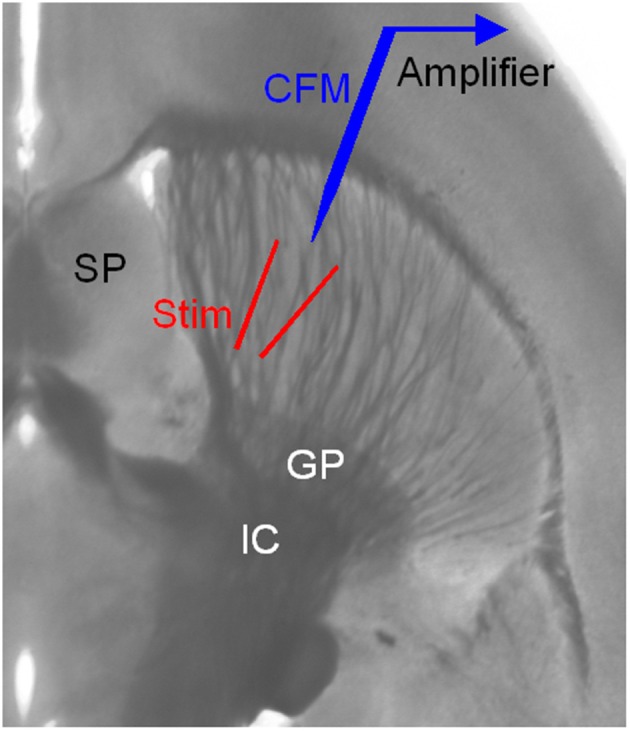
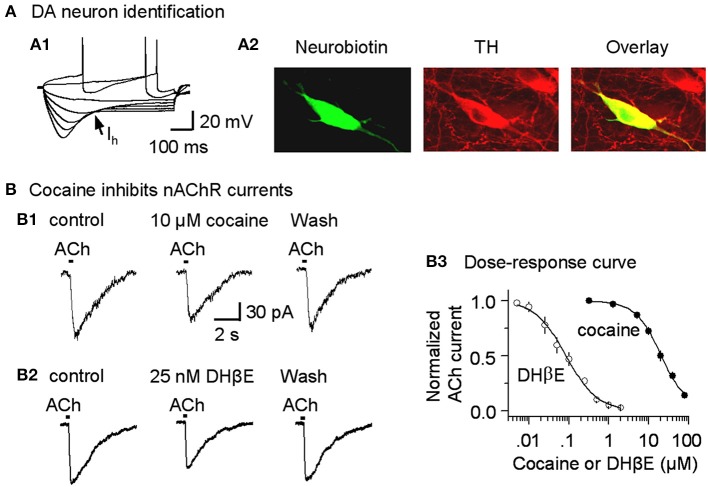
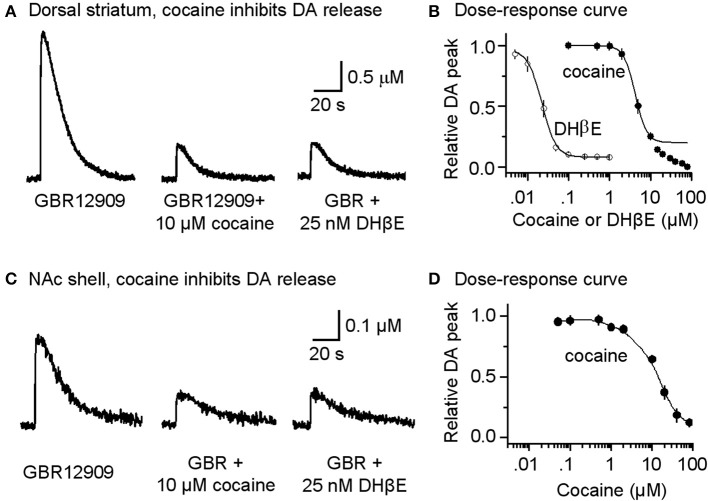
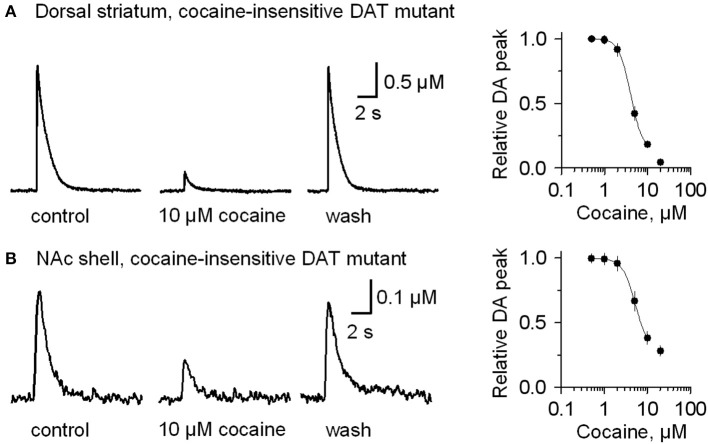
References
-
- Alkondon M., Albuquerque E. X. (1993). Diversity of nicotinic acetylcholine receptors in rat hippocampal neurons. I. Pharmacological and functional evidence for distinct structural subtypes. J. Pharmacol. Exp. Ther. 265, 1455–1473 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources