

The DExH box helicase domain of spindle-E is necessary for retrotransposon silencing and axial patterning during Drosophila oogenesis
- PMID: 25239103
- PMCID: PMC4232550
- DOI: 10.1534/g3.114.014332
The DExH box helicase domain of spindle-E is necessary for retrotransposon silencing and axial patterning during Drosophila oogenesis
Abstract
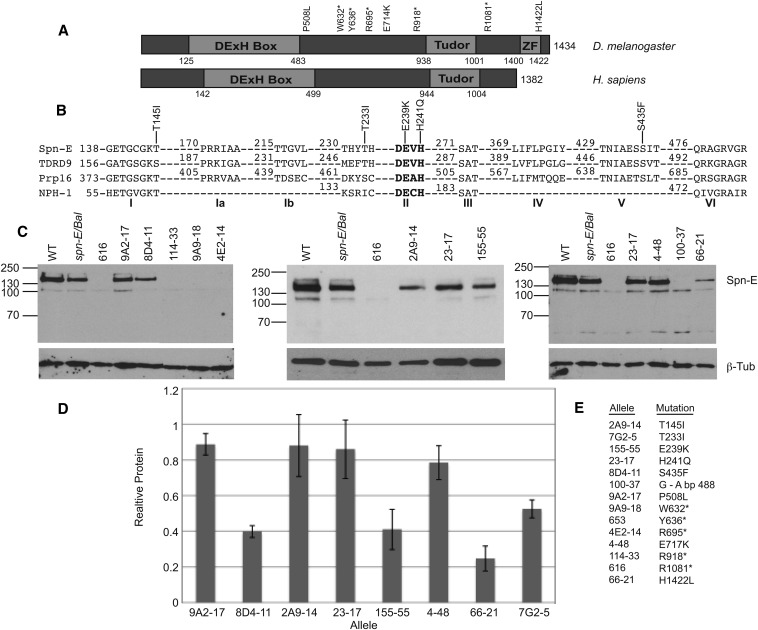
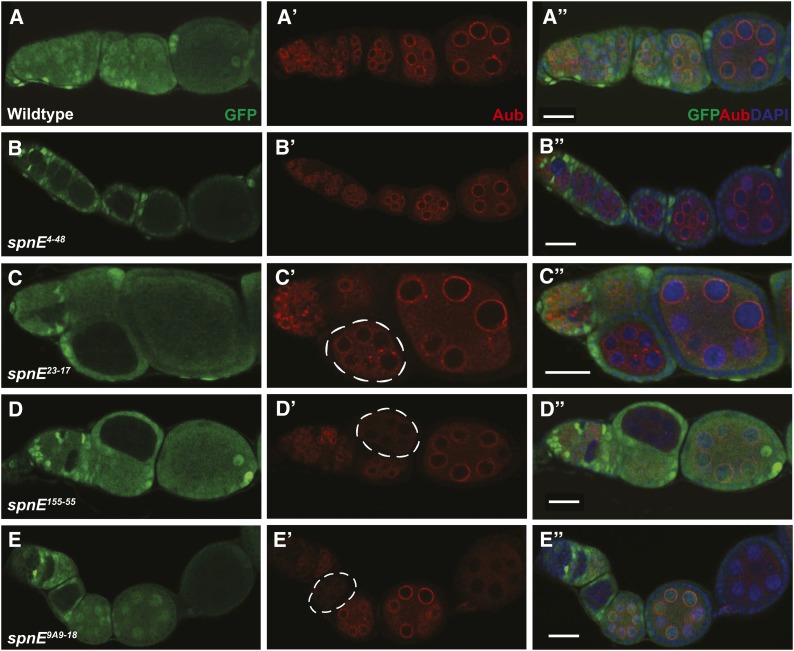
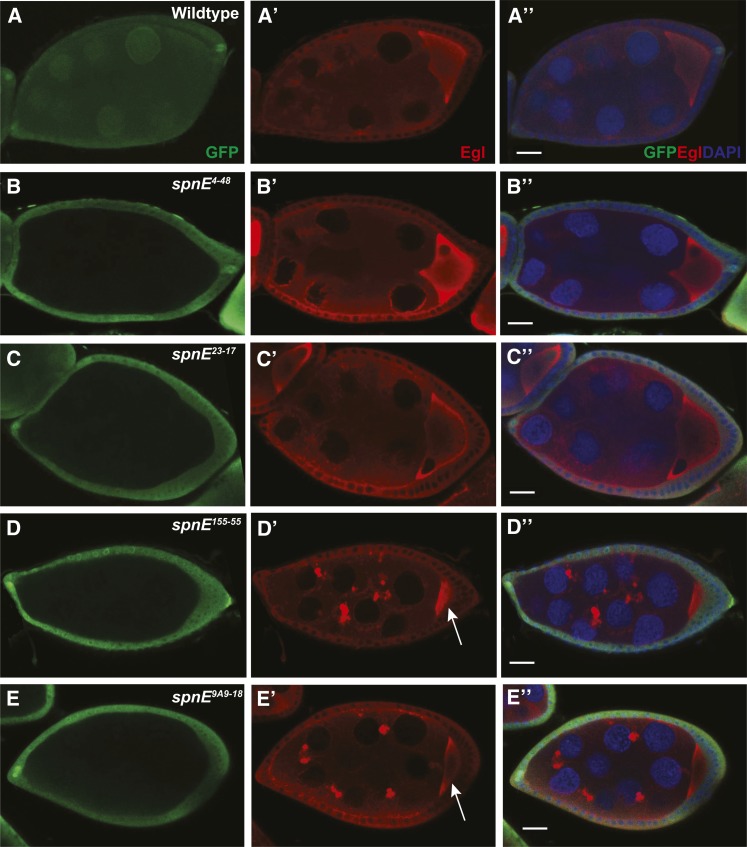
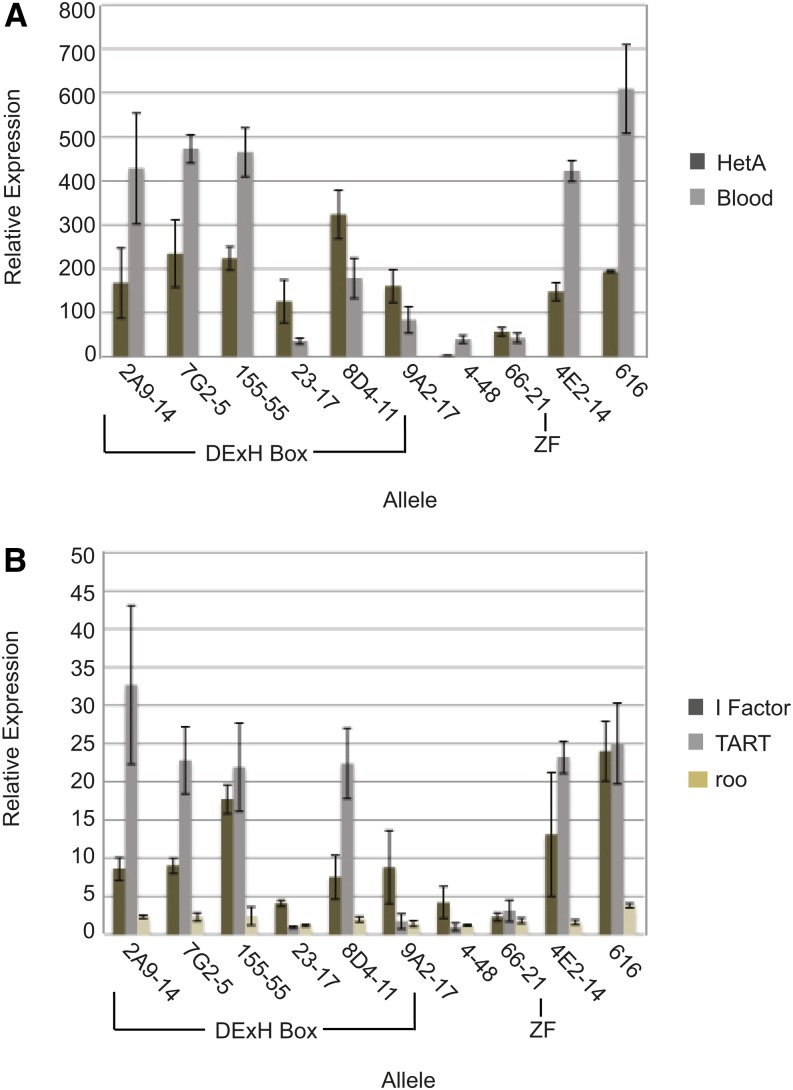
Transposable selfish genetic elements have the potential to cause debilitating mutations as they replicate and reinsert within the genome. Therefore, it is critical to keep the cellular levels of these elements low. This is especially true in the germline where these mutations could affect the viability of the next generation. A class of small noncoding RNAs, the Piwi-associated RNAs, is responsible for silencing transposable elements in the germline of most organisms. Several proteins have been identified as playing essential roles in piRNA generation and transposon silencing. However, for the most part their function in piRNA generation is currently unknown. One of these proteins is the Drosophila melanogaster DExH box/Tudor domain protein Spindle-E, whose activity is necessary for the generation of most germline piRNAs. In this study we molecularly and phenotypically characterized 14 previously identified spindle-E alleles. Of the alleles that express detectable Spindle-E protein, we found that five had mutations in the DExH box domain. Additionally, we found that processes that depend on piRNA function, including Aubergine localization, Dynein motor movement, and retrotransposon silencing, were severely disrupted in alleles with DExH box domain mutations. The phenotype of many of these alleles is as severe as the strongest spindle-E phenotype, whereas alleles with mutations in other regions of Spindle-E did not affect these processes as much. From these data we conclude that the DExH box domain of Spindle-E is necessary for its function in the piRNA pathway and retrotransposon silencing.
Keywords: Drosophila; RNA helicase; embryonic patterning; oogenesis; piRNA; retrotransposon.
Copyright © 2014 Ott et al.
Figures
Similar articles
-
[The interplay of transposon silencing genes in the Drosophila melanogaster germline].Mol Biol (Mosk). 2011 Jul-Aug;45(4):633-41. Mol Biol (Mosk). 2011. PMID: 21954595 Russian.
-
The Tudor domain protein Tapas, a homolog of the vertebrate Tdrd7, functions in the piRNA pathway to regulate retrotransposons in germline of Drosophila melanogaster.BMC Biol. 2014 Oct 6;12:61. doi: 10.1186/s12915-014-0061-9. BMC Biol. 2014. PMID: 25287931 Free PMC article.
-
RNA helicase Spn-E is required to maintain Aub and AGO3 protein levels for piRNA silencing in the germline of Drosophila.Eur J Cell Biol. 2016 Sep;95(9):311-22. doi: 10.1016/j.ejcb.2016.06.001. Epub 2016 Jun 6. Eur J Cell Biol. 2016. PMID: 27320195
-
piRNA pathway and the potential processing site, the nuage, in the Drosophila germline.Dev Growth Differ. 2012 Jan;54(1):66-77. doi: 10.1111/j.1440-169x.2011.01316.x. Dev Growth Differ. 2012. PMID: 23741748 Review.
-
piRNAs, transposon silencing, and Drosophila germline development.J Cell Biol. 2010 Nov 29;191(5):905-13. doi: 10.1083/jcb.201006034. J Cell Biol. 2010. PMID: 21115802 Free PMC article. Review.
Cited by
-
The dual role of Spn-E in supporting heterotypic ping-pong piRNA amplification in silkworms.EMBO Rep. 2024 May;25(5):2239-2257. doi: 10.1038/s44319-024-00137-2. Epub 2024 Apr 17. EMBO Rep. 2024. PMID: 38632376 Free PMC article.
-
Tejas functions as a core component in nuage assembly and precursor processing in Drosophila piRNA biogenesis.J Cell Biol. 2023 Oct 2;222(10):e202303125. doi: 10.1083/jcb.202303125. Epub 2023 Aug 9. J Cell Biol. 2023. PMID: 37555815 Free PMC article.
-
Spindle-E Acts Antivirally Against Alphaviruses in Mosquito Cells.Viruses. 2018 Feb 18;10(2):88. doi: 10.3390/v10020088. Viruses. 2018. PMID: 29463033 Free PMC article.
-
Epigenetic roles of PIWI proteins and piRNAs in colorectal cancer.Cancer Cell Int. 2021 Jun 30;21(1):328. doi: 10.1186/s12935-021-02034-3. Cancer Cell Int. 2021. PMID: 34193172 Free PMC article. Review.
-
Suppression of Transposable Elements in Leukemic Stem Cells.Sci Rep. 2017 Aug 1;7(1):7029. doi: 10.1038/s41598-017-07356-9. Sci Rep. 2017. PMID: 28765607 Free PMC article.
References
-
- Brennecke J., Aravin A. A., Stark A., Dus M., Kellis M., et al. , 2007. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128: 1089–1103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases