

Insulin is involved in transcriptional regulation of NKCC and the CFTR Cl(-) channel through PI3K activation and ERK inactivation in renal epithelial cells
- PMID: 25239597
- PMCID: PMC10717268
- DOI: 10.1007/s12576-014-0338-3
Insulin is involved in transcriptional regulation of NKCC and the CFTR Cl(-) channel through PI3K activation and ERK inactivation in renal epithelial cells
Abstract
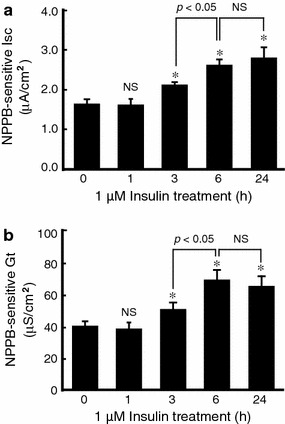
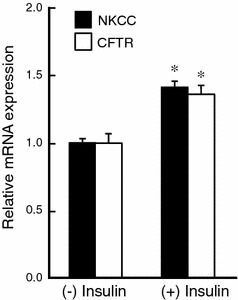
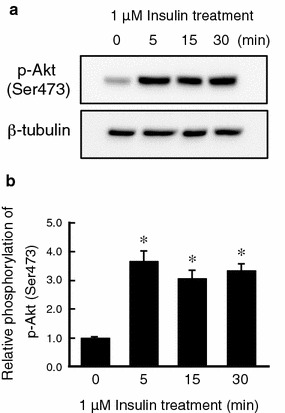
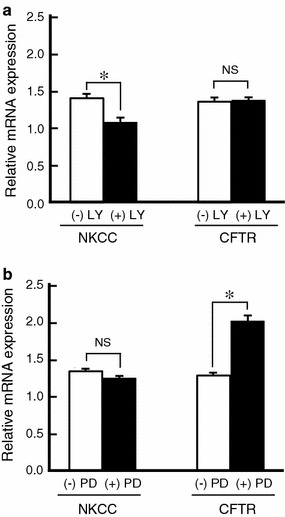
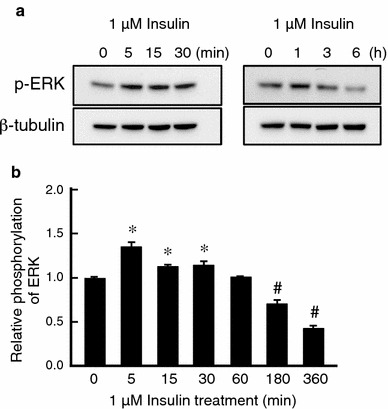
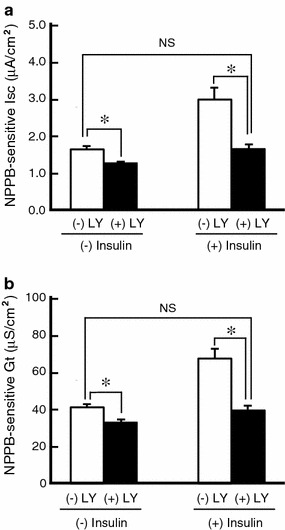
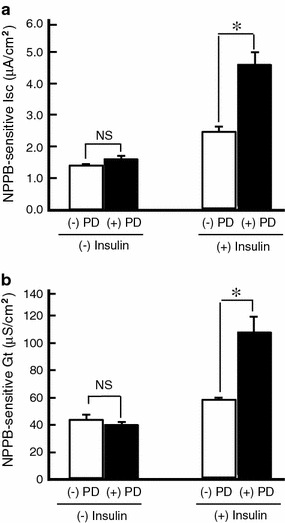
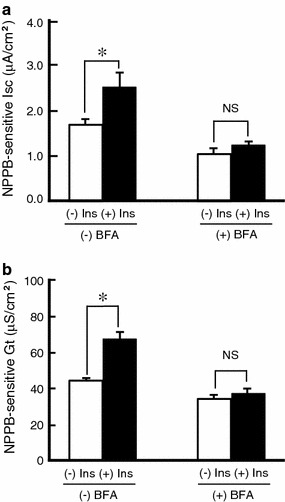
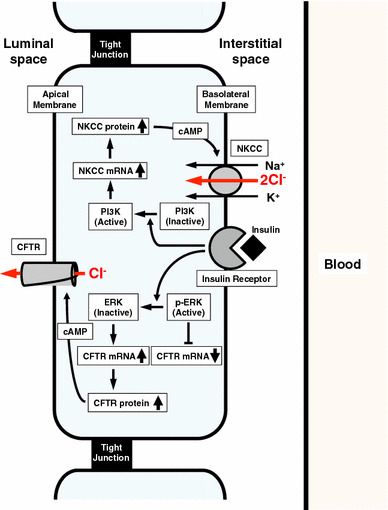
It is is well known that insulin stimulates glucose transport and epithelial Na(+) channel (ENaC)-mediated Na(+) reabsorption; however, the action of insulin on Cl(-) secretion is not fully understood. In this study, we investigated the action of insulin on Na(+)-K(+)-2Cl(-) cotransporter (NKCC)-mediated Cl(-) secretion in epithelial A6 cells. Interestingly, insulin treatment remarkably enhanced the forskolin-stimulated Cl(-) secretion associated with an increase in apical Cl(-) conductance by upregulating mRNA expression of both CFTR and NKCC, although insulin treatment alone had no effect on the basal Cl(-) secretion or apical Cl(-) conductance without forskolin application. We next elucidated a role of phosphoinositide 3-kinase (PI3K) in the insulin-induced enhancement of the Cl(-) secretion, since insulin actually activated PI3K, resulting in activation of Akt, a downstream molecule of PI3K. LY294002 (a PI3K inhibitor) reduced the Cl(-) secretion by suppressing mRNA expression of NKCC, whereas insulin still had a stimulatory action on mRNA expression of CFTR even in the presence of LY294002. On the other hand, we found that a MEK inhibitor (PD98059) further enhanced the insulin-stimulated CFTR mRNA expression and the Cl(-) secretion in forskolin-stimulated A6 cells and that insulin induced slight, transient activation of ERK followed by significant inactivation of ERK. These observations suggest that: (1) insulin respectively upregulates mRNA expression of NKCC and CFTR through activation of PI3K and inactivation of ERK; (2) insulin signals on mRNA expression of NKCC and CFTR are not enough to stimulate transepithelial Cl(-) secretion, but enhance the stimulatory action of cAMP on transepithelial Cl(-) secretion.
Figures
References
-
- Marunaka Y (2014) Characteristics and pharmacological regulation of epithelial Na+ channel (ENaC) and epithelial Na+ transport. J Pharmacol Sci. (in press). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
