

Tasks for inhibitory interneurons in intact brain circuits
- PMID: 25239808
- PMCID: PMC4254329
- DOI: 10.1016/j.neuropharm.2014.09.011
Tasks for inhibitory interneurons in intact brain circuits
Abstract
Synaptic inhibition, brought about by a rich variety of interneuron types, counters excitation, modulates the gain, timing, tuning, bursting properties of principal cell firing, and exerts selective filtering of synaptic excitation. At the network level, it allows for coordinating transient interactions among the principal cells to form cooperative assemblies for efficient transmission of information and routing of excitatory activity across networks, typically in the form of brain oscillations. Recent techniques based on targeted expression of neuronal activity modulators, such as optogenetics, allow physiological identification and perturbation of specific interneuron subtypes in the intact brain. Combined with large-scale recordings or imaging techniques, these approaches facilitate our understanding of the multiple roles of inhibitory interneurons in shaping circuit functions.
Keywords: Circuits; Inhibition; Interneurons; Optogenetics; Oscillations; Pharmacogenetics.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
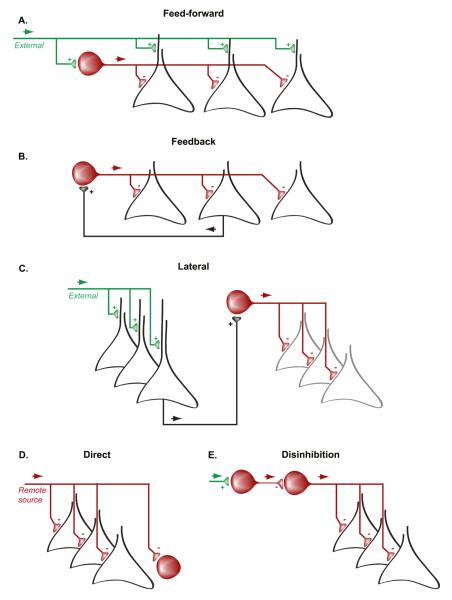

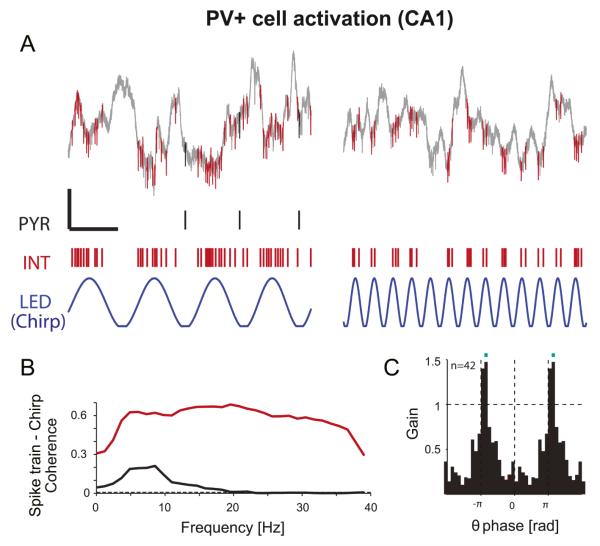
References
-
- Alonso A, Kohler C. Evidence for separate projections of hippocampal pyramidal and nonpyramidal neurons to different parts of the septum in the rat brain. Neuroscience letters. 1982;31:209–214. - PubMed
-
- Alonso A, Kohler C. A study of the reciprocal connections between the septum and the entorhinal area using anterograde and retrograde axonal transport methods in the rat brain. The Journal of comparative neurology. 1984;225:327–343. - PubMed
-
- Alonso M, Lepousez G, Sebastien W, Bardy C, Gabellec MM, Torquet N, Lledo PM. Activation of adult-born neurons facilitates learning and memory. Nat Neurosci. 2012;15:897–904. - PubMed
-
- Anderson JS, Carandini M, Ferster D. Orientation tuning of input conductance, excitation, and inhibition in cat primary visual cortex. J Neurophysiol. 2000;84:909–926. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
