

Trim58 degrades Dynein and regulates terminal erythropoiesis
- PMID: 25241935
- PMCID: PMC4203341
- DOI: 10.1016/j.devcel.2014.07.021
Trim58 degrades Dynein and regulates terminal erythropoiesis
Abstract
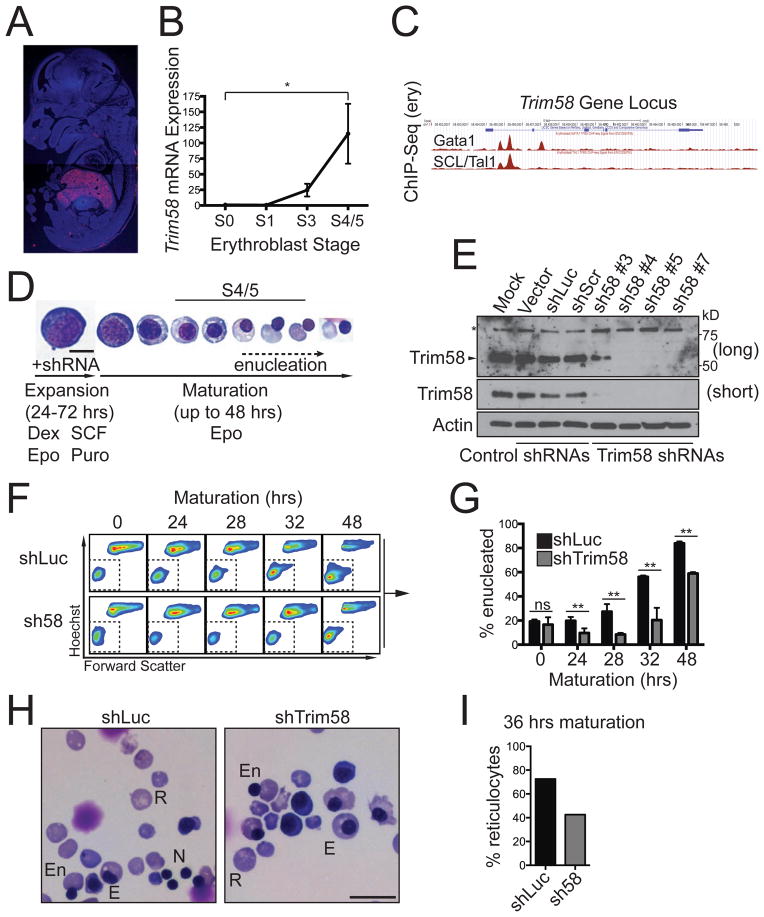
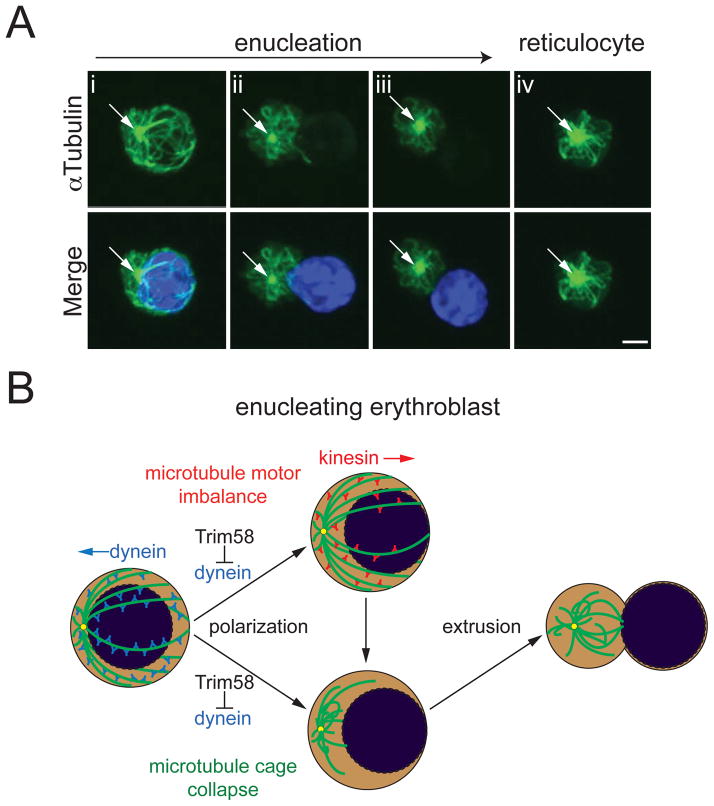
TRIM58 is an E3 ubiquitin ligase superfamily member implicated by genome-wide association studies to regulate human erythrocyte traits. Here, we show that Trim58 expression is induced during late erythropoiesis and that its depletion by small hairpin RNAs (shRNAs) inhibits the maturation of late-stage nucleated erythroblasts to anucleate reticulocytes. Imaging flow cytometry studies demonstrate that Trim58 regulates polarization and/or extrusion of erythroblast nuclei. In vitro, Trim58 directly binds and ubiquitinates the intermediate chain of the microtubule motor dynein. In cells, Trim58 stimulates proteasome-dependent degradation of the dynein holoprotein complex. During erythropoiesis, Trim58 expression, dynein loss, and enucleation occur concomitantly, and all are inhibited by Trim58 shRNAs. Dynein regulates nuclear positioning and microtubule organization, both of which undergo dramatic changes during erythroblast enucleation. Thus, we propose that Trim58 promotes this process by eliminating dynein. Our findings identify an erythroid-specific regulator of enucleation and elucidate a previously unrecognized mechanism for controlling dynein activity.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




References
-
- Chen CY, Pajak L, Tamburlin J, Bofinger D, Koury ST. The effect of proteasome inhibitors on mammalian erythroid terminal differentiation. Exp Hematol. 2002;30:634–639. - PubMed
-
- Dolznig H, Bartunek P, Nasmyth K, Mullner EW, Beug H. Terminal differentiation of normal chicken erythroid progenitors: shortening of G1 correlates with loss of D-cyclin/cdk4 expression and altered cell size control. Cell Growth Differ. 1995;6:1341–1352. - PubMed
-
- Fabrini R, De Luca A, Stella L, Mei G, Orioni B, Ciccone S, Federici G, Lo Bello M, Ricci G. Monomer-dimer equilibrium in glutathione transferases: a critical re-examination. Biochemistry. 2009;48:10473–10482. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 HL099656/HL/NHLBI NIH HHS/United States
- P30 DK090969/DK/NIDDK NIH HHS/United States
- F30 DK102291/DK/NIDDK NIH HHS/United States
- R56 DK061692/DK/NIDDK NIH HHS/United States
- T32 GM007170/GM/NIGMS NIH HHS/United States
- T32 GM008216/GM/NIGMS NIH HHS/United States
- R01 DK061692/DK/NIDDK NIH HHS/United States
- R01 GM048661/GM/NIGMS NIH HHS/United States
- U01 HL099993/HL/NHLBI NIH HHS/United States
- P01 HL110860/HL/NHLBI NIH HHS/United States
- DK61692/DK/NIDDK NIH HHS/United States
- GM007170/GM/NIGMS NIH HHS/United States
- T32 HL007439/HL/NHLBI NIH HHS/United States
- HL007439/HL/NHLBI NIH HHS/United States
- P01 HL064190/HL/NHLBI NIH HHS/United States
- P30DK090969/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
