

Single rodent mesohabenular axons release glutamate and GABA
- PMID: 25242304
- PMCID: PMC4843828
- DOI: 10.1038/nn.3823
Single rodent mesohabenular axons release glutamate and GABA
Abstract
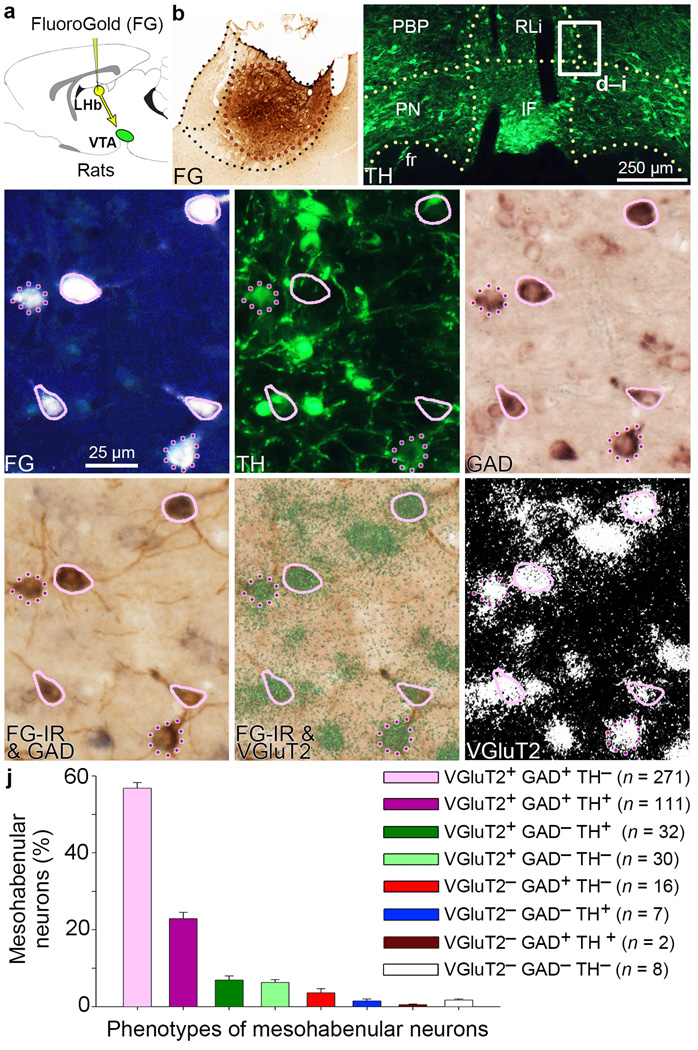
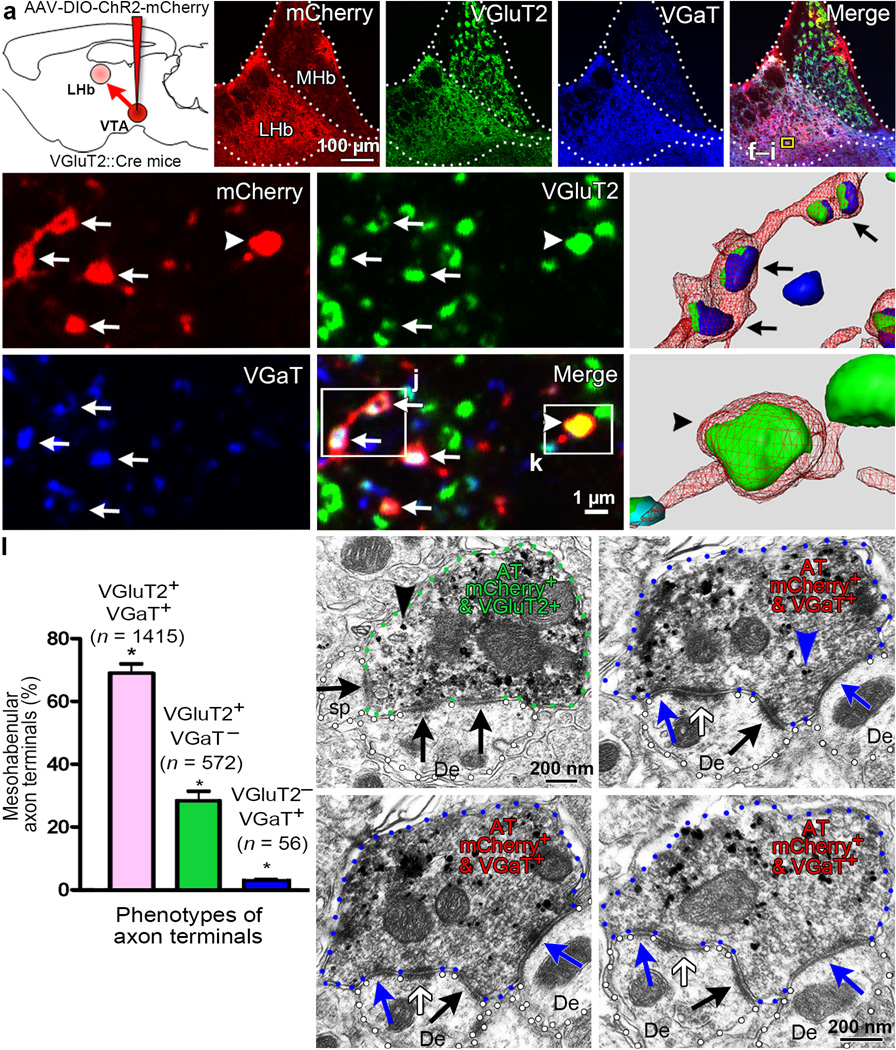
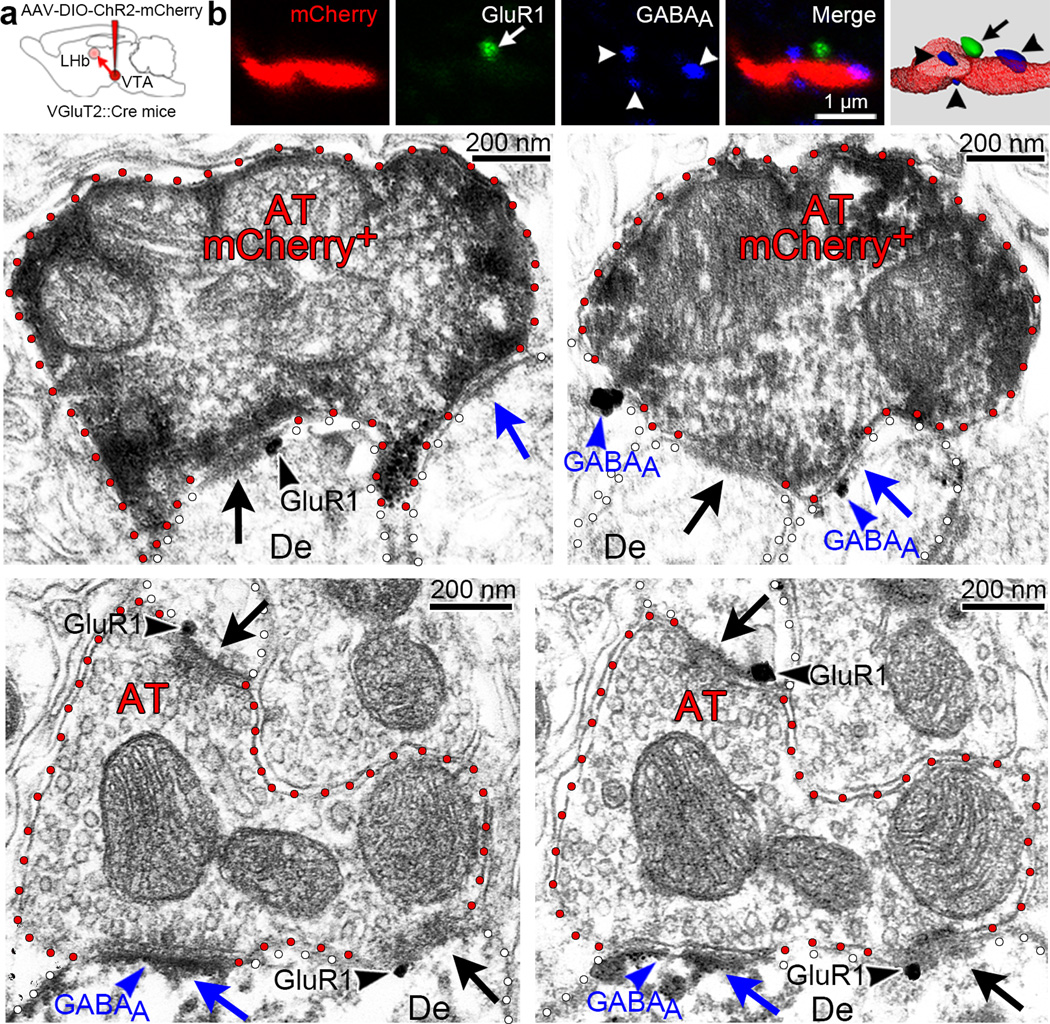
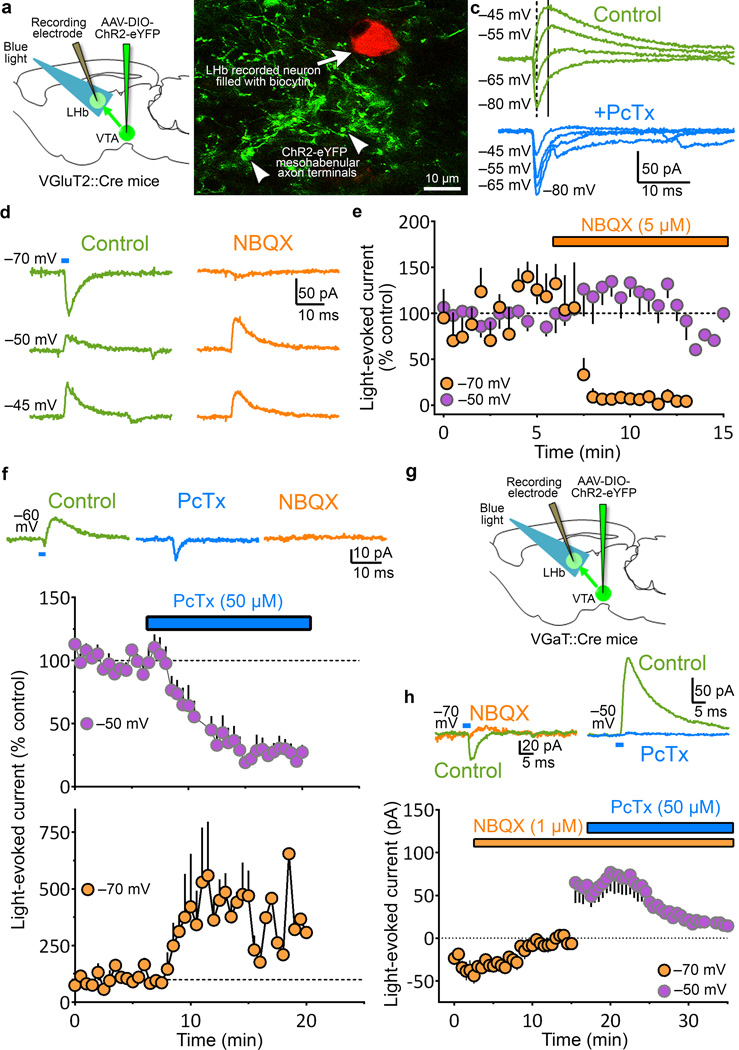
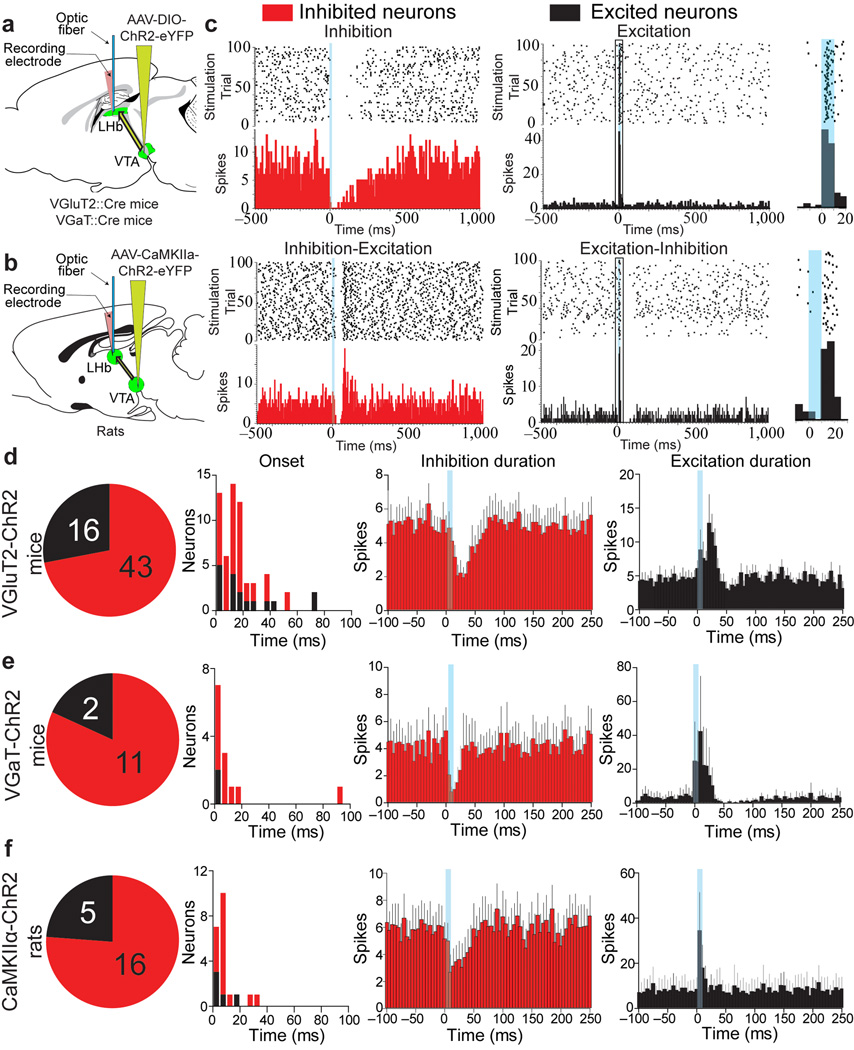
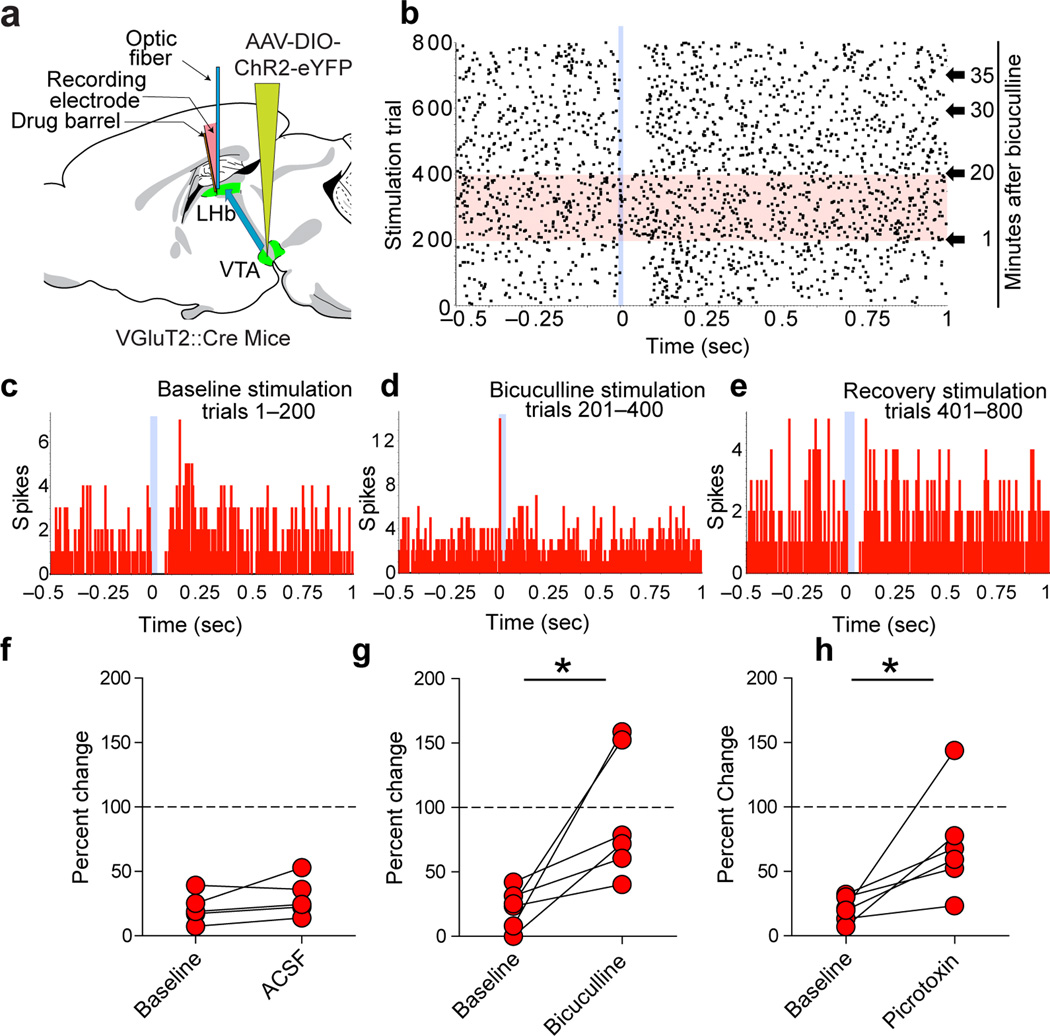
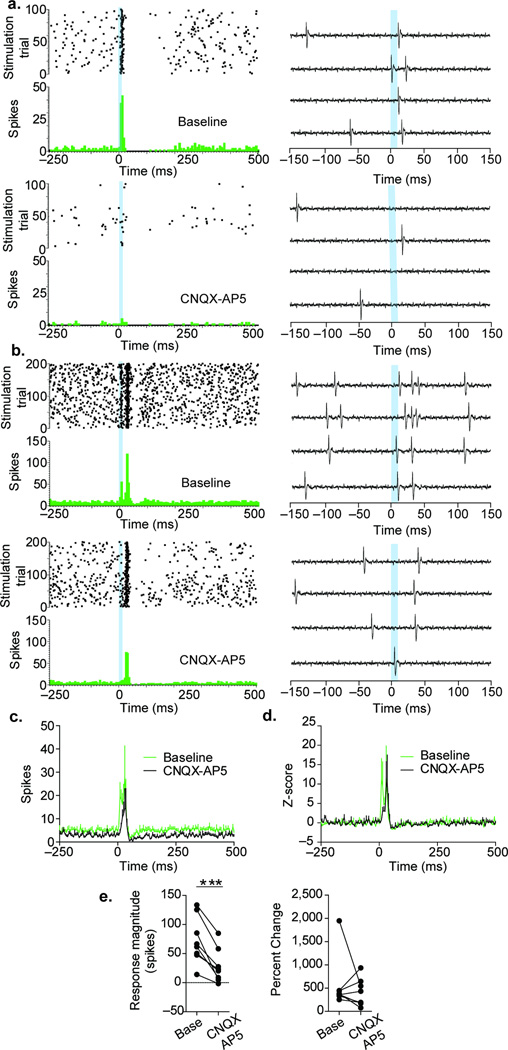
The lateral habenula (LHb) is involved in reward, aversion, addiction and depression through descending interactions with several brain structures, including the ventral tegmental area (VTA). The VTA provides reciprocal inputs to LHb, but their actions are unclear. Here we show that the majority of rat and mouse VTA neurons innervating LHb coexpress markers for both glutamate signaling (vesicular glutamate transporter 2; VGluT2) and GABA signaling (glutamic acid decarboxylase; GAD, and vesicular GABA transporter; VGaT). A single axon from these mesohabenular neurons coexpresses VGluT2 protein and VGaT protein and, surprisingly, establishes symmetric and asymmetric synapses on LHb neurons. In LHb slices, light activation of mesohabenular fibers expressing channelrhodopsin2 driven by VGluT2 (Slc17a6) or VGaT (Slc32a1) promoters elicits release of both glutamate and GABA onto single LHb neurons. In vivo light activation of mesohabenular terminals inhibits or excites LHb neurons. Our findings reveal an unanticipated type of VTA neuron that cotransmits glutamate and GABA and provides the majority of mesohabenular inputs.
Conflict of interest statement
The authors declare that they do not have any conflicts of interest (financial or otherwise) related to the data presented in this manuscript.
Figures
Comment in
-
Neurotransmission: Transmission takes two.Nat Rev Neurosci. 2014 Nov;15(11):700-1. doi: 10.1038/nrn3848. Epub 2014 Oct 15. Nat Rev Neurosci. 2014. PMID: 25315393 No abstract available.
-
Bilingual neurons release glutamate and GABA.Nat Neurosci. 2014 Nov;17(11):1432-4. doi: 10.1038/nn.3840. Nat Neurosci. 2014. PMID: 25349905 No abstract available.
References
-
- Matsumoto M, Hikosaka O. Lateral habenula as a source of negative reward signals in dopamine neurons. Nature. 2007;447(7148):1111–1115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
