

The novel C24D synthetic polypeptide inhibits binding of placenta immunosuppressive ferritin to human T cells and elicits anti-breast cancer immunity in vitro and in vivo
- PMID: 25246274
- PMCID: PMC4235009
- DOI: 10.1016/j.neo.2014.08.005
The novel C24D synthetic polypeptide inhibits binding of placenta immunosuppressive ferritin to human T cells and elicits anti-breast cancer immunity in vitro and in vivo
Abstract
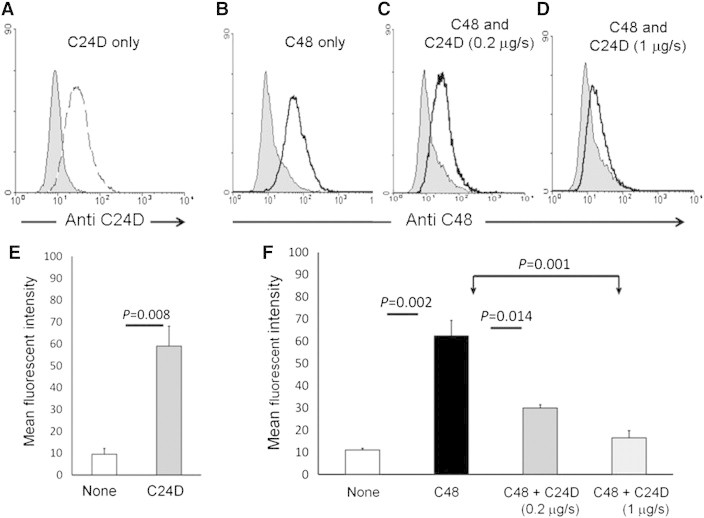
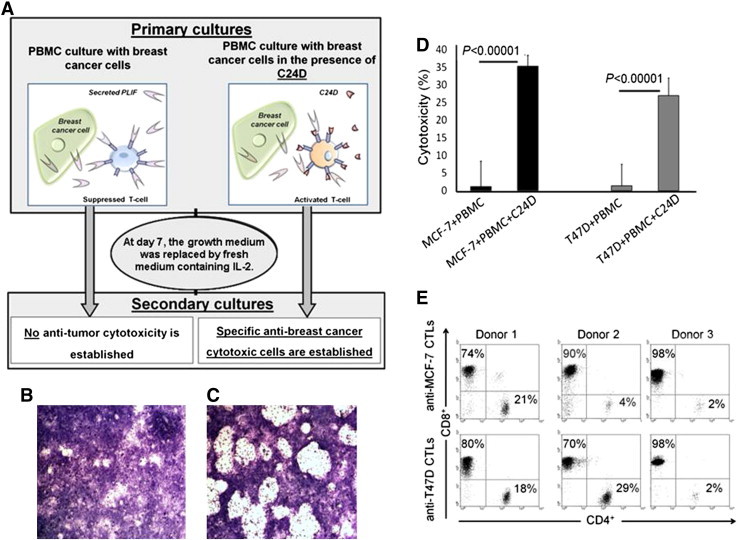
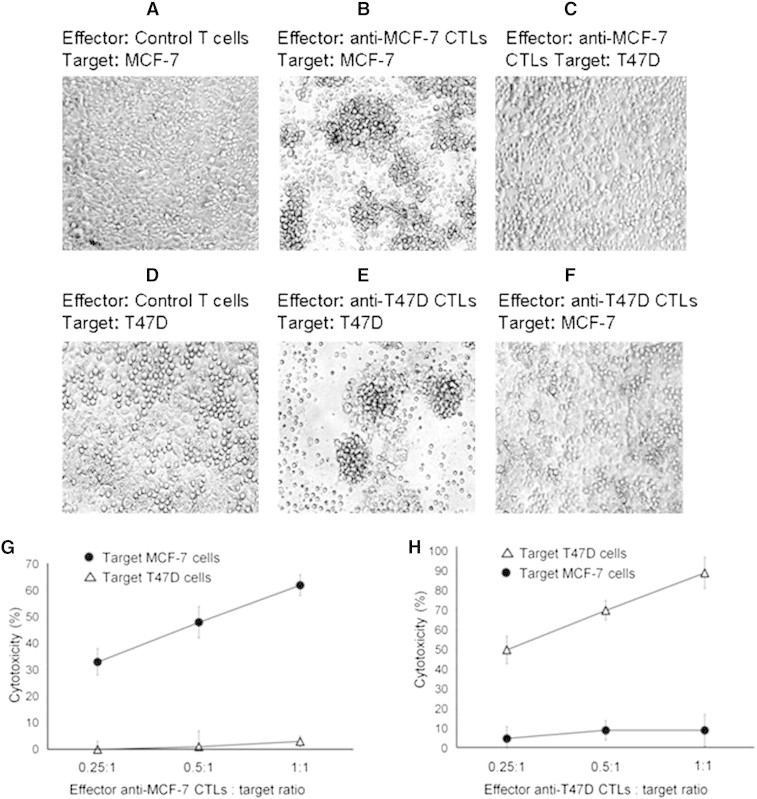
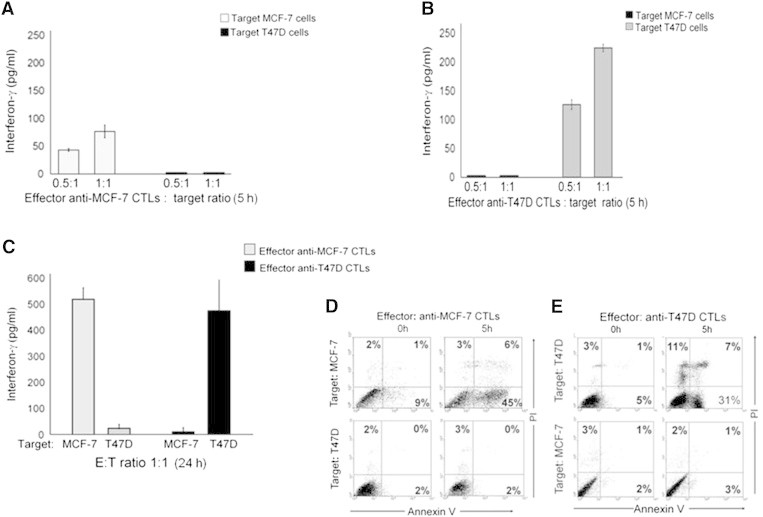
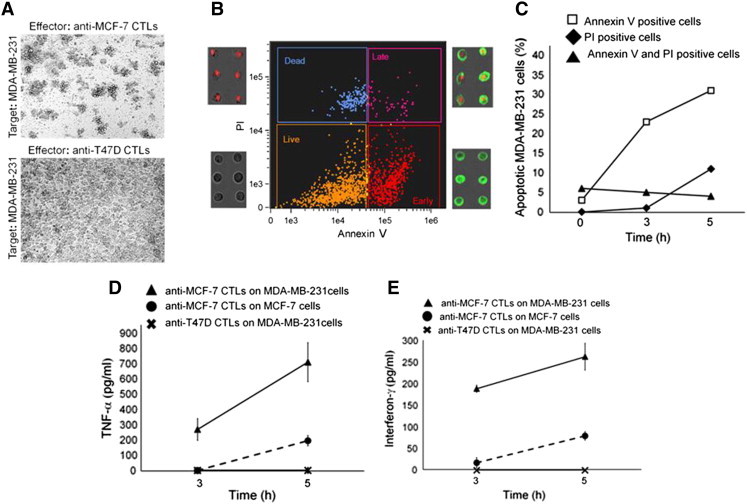
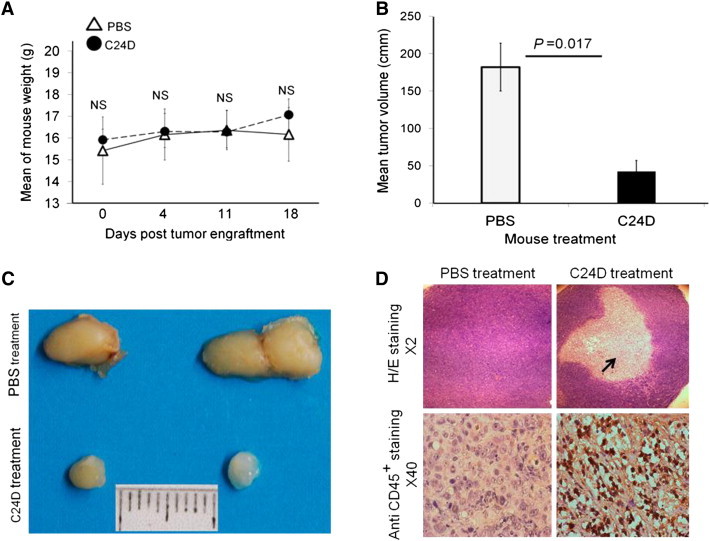
Immune tolerance mechanisms supporting normal human pregnancy are exploited by breast cancer and other malignancies. We cloned from human placenta and breast cancer cells the novel human immunomodulator named placenta immunosuppressive ferritin (PLIF). PLIF is composed of a ferritin heavy chain-like domain and a novel cytokine-like domain, named C48. Both intact PLIF and C48 inhibit T cell proliferation. Blocking PLIF by specific antibodies in a tolerant breast cancer model in nude mice resulted in tumor cell apoptosis and rejection. This prompted us to study active immune preventive strategies targeting PLIF activity. Currently, we report on the design and synthesis of the novel C24D polypeptide, which inhibits the binding of PLIF to T cells and therefore inhibits the immune suppressive effect of PLIF. The effect of C24D on the generation of anti-breast cancer cytotoxic T lymphocytes (CTLs) was studied in vitro in cultures of MCF-7 (HLA-A2(+)) or T47D (HLA-A2(-)) breast cancer cells incubated with peripheral blood mononuclear cells (PBMCs) from healthy blood donors. We found that C24D treatment exclusively induced development of CTLs. On reactivation by their specific target cells, the CTLs secreted interferon-γ and induced target apoptosis. Anti-MCF-7 CTLs were cross-cytotoxic to MDA-MB-231 (HLA-A2(+)) triple-negative breast cancer but not to T47D. Moreover, C24D treatment in vivo inhibited the growth of MCF-7 tumors engrafted in immune-compromised nude mice transfused with naïve allogeneic human PBMCs. Our results demonstrate that C24D treatment breakdown breast cancer induced tolerance enabling the initiation of effective anti-tumor immune response.
Copyright © 2014 Neoplasia Press, Inc. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
A novel role for an old target: CD45 for breast cancer immunotherapy.Oncoimmunology. 2021 May 25;10(1):1929725. doi: 10.1080/2162402X.2021.1929725. Oncoimmunology. 2021. PMID: 34104545 Free PMC article.
-
Antibodies to placental immunoregulatory ferritin with transfer of polyclonal lymphocytes arrest MCF-7 human breast cancer growth in a nude mouse model.Neoplasia. 2007 Jun;9(6):487-94. doi: 10.1593/neo.07259. Neoplasia. 2007. PMID: 17603631 Free PMC article.
-
PLIF, a novel human ferritin subunit from placenta with immunosuppressive activity.J Biol Chem. 2002 Apr 12;277(15):12901-5. doi: 10.1074/jbc.M200956200. Epub 2002 Jan 30. J Biol Chem. 2002. PMID: 11821435
-
Identification of a novel HLA-A2-restricted cytotoxic T lymphocyte epitope from cancer-testis antigen PLAC1 in breast cancer.Amino Acids. 2012 Jun;42(6):2257-65. doi: 10.1007/s00726-011-0966-3. Epub 2011 Jun 28. Amino Acids. 2012. PMID: 21710262
-
PLIF induces IL-10 production in monocytes: a calmodulin-p38 mitogen-activated protein kinase-dependent pathway.FASEB J. 2003 May;17(8):955-7. doi: 10.1096/fj.02-0960fje. Epub 2003 Mar 28. FASEB J. 2003. PMID: 12670872
Cited by
-
Pregnancy complications and subsequent breast cancer risk in the mother: a Nordic population-based case-control study.Int J Cancer. 2018 Oct 15;143(8):1904-1913. doi: 10.1002/ijc.31600. Epub 2018 Aug 10. Int J Cancer. 2018. PMID: 29752724 Free PMC article.
-
A novel role for an old target: CD45 for breast cancer immunotherapy.Oncoimmunology. 2021 May 25;10(1):1929725. doi: 10.1080/2162402X.2021.1929725. Oncoimmunology. 2021. PMID: 34104545 Free PMC article.
-
Downregulation of CD45 Signaling in COVID-19 Patients Is Reversed by C24D, a Novel CD45 Targeting Peptide.Front Med (Lausanne). 2021 Aug 3;8:675963. doi: 10.3389/fmed.2021.675963. eCollection 2021. Front Med (Lausanne). 2021. PMID: 34414199 Free PMC article.
-
Facts and prospects of peptide in targeted therapy and immune regulation against triple-negative breast cancer.Front Immunol. 2023 Aug 25;14:1255820. doi: 10.3389/fimmu.2023.1255820. eCollection 2023. Front Immunol. 2023. PMID: 37691919 Free PMC article. Review.
References
-
- Kirkwood J. Cancer immunotherapy: the interferon-alpha experience. Semin Oncol. 2002;29:18–26. - PubMed
-
- Boyiadzis M., Foon K.A. Approved monoclonal antibodies for cancer therapy. Expert Opin Biol Ther. 2008;8:1151–1158. - PubMed
-
- Nimjee S.M., Rusconi C.P., Sullenger B.A. Aptamers: an emerging class of therapeutics. Annu Rev Med. 2005;56:555–583. - PubMed
-
- van der Burg S.H., Bijker M.S., Welters M.J., Offringa R., Melief C.J. Improved peptide vaccine strategies, creating synthetic artificial infections to maximize immune efficacy. Adv Drug Deliv Rev. 2006;58:916–930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
