

Second transmembrane helix (M2) and long range coupling in Ca²⁺-ATPase
- PMID: 25246522
- PMCID: PMC4223325
- DOI: 10.1074/jbc.M114.584086
Second transmembrane helix (M2) and long range coupling in Ca²⁺-ATPase
Abstract
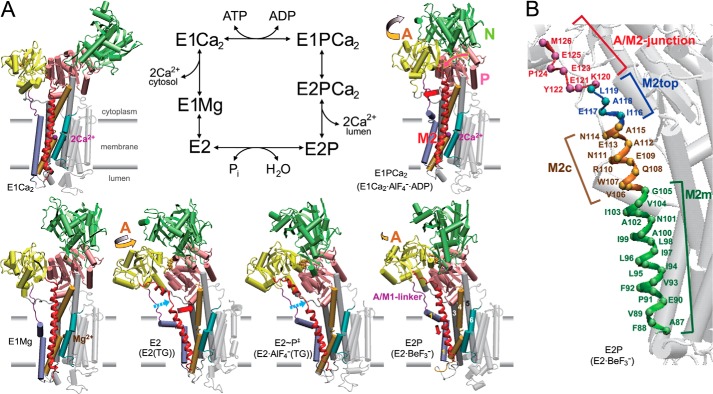
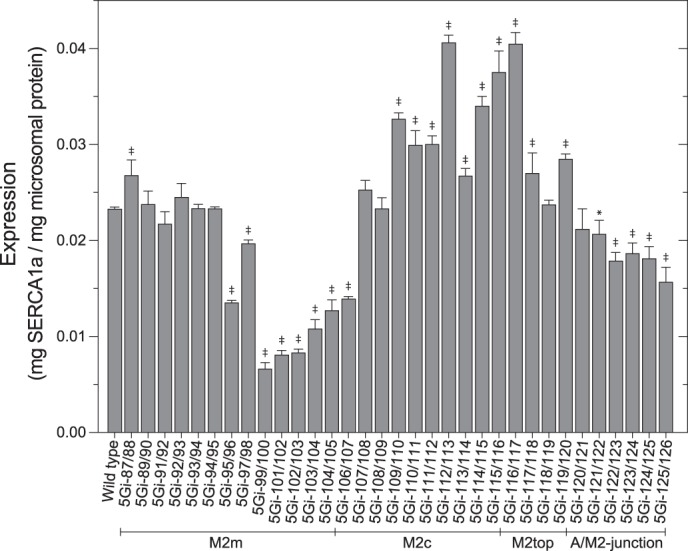
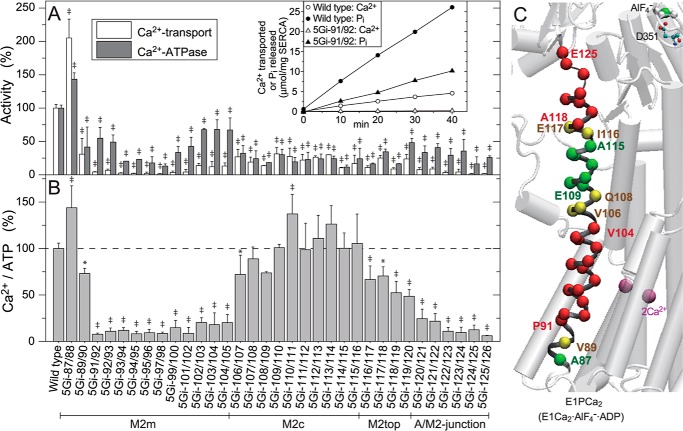
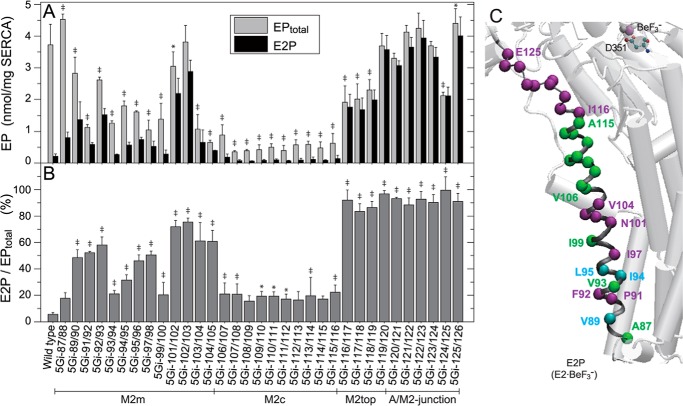
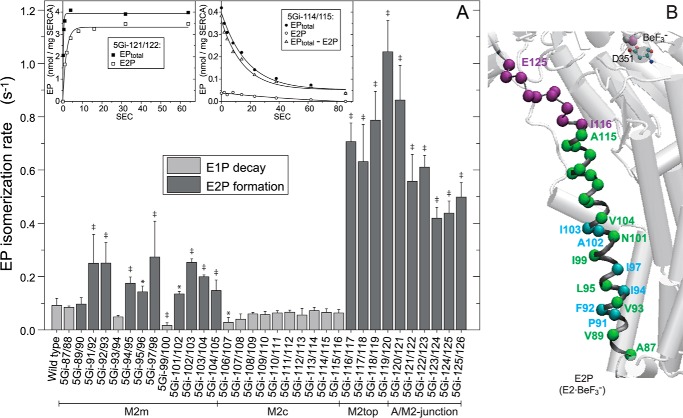
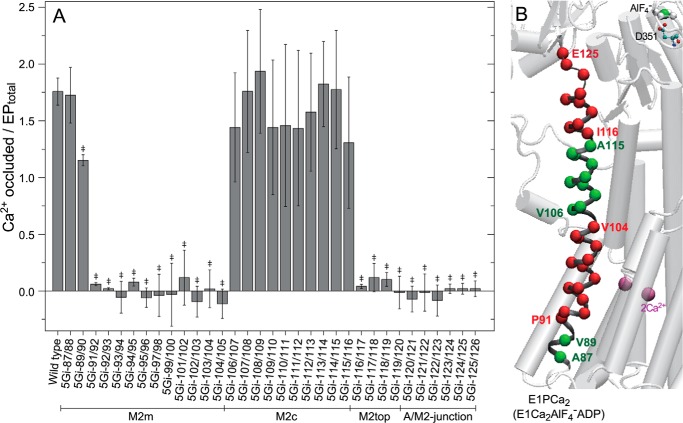
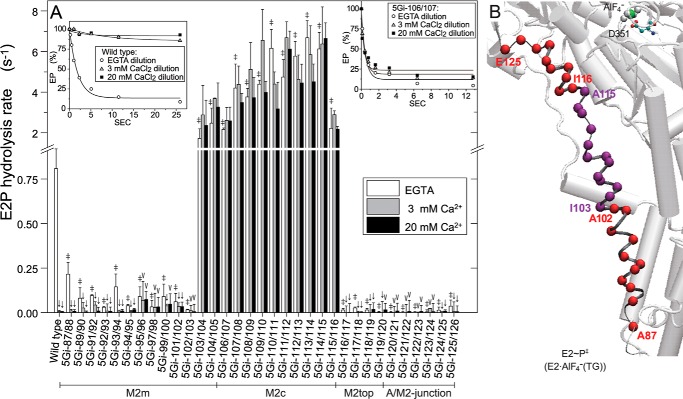
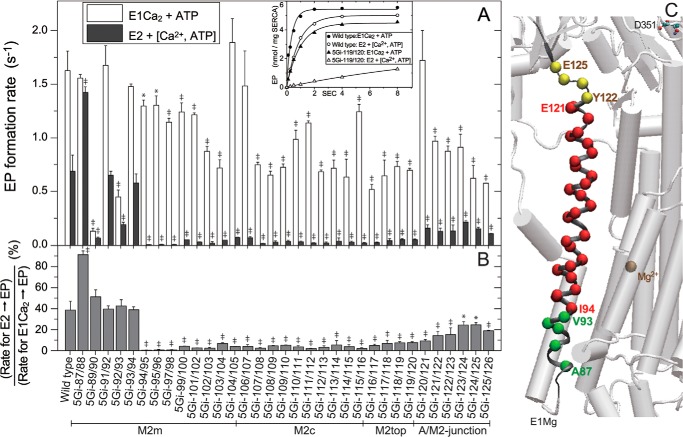
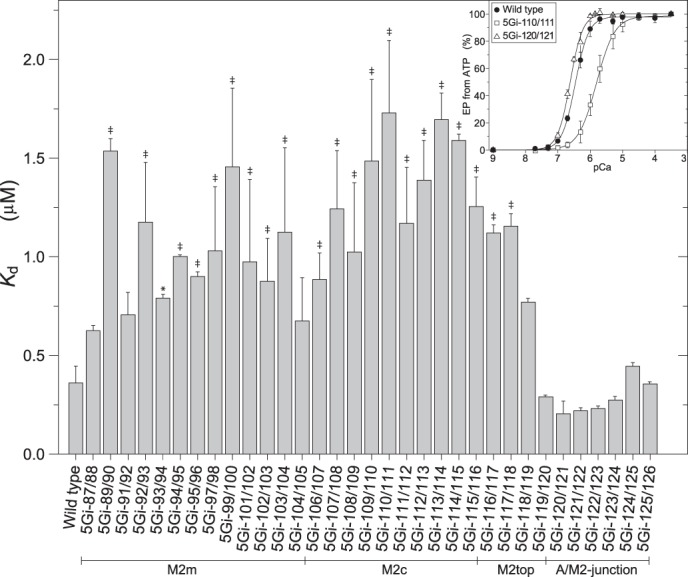
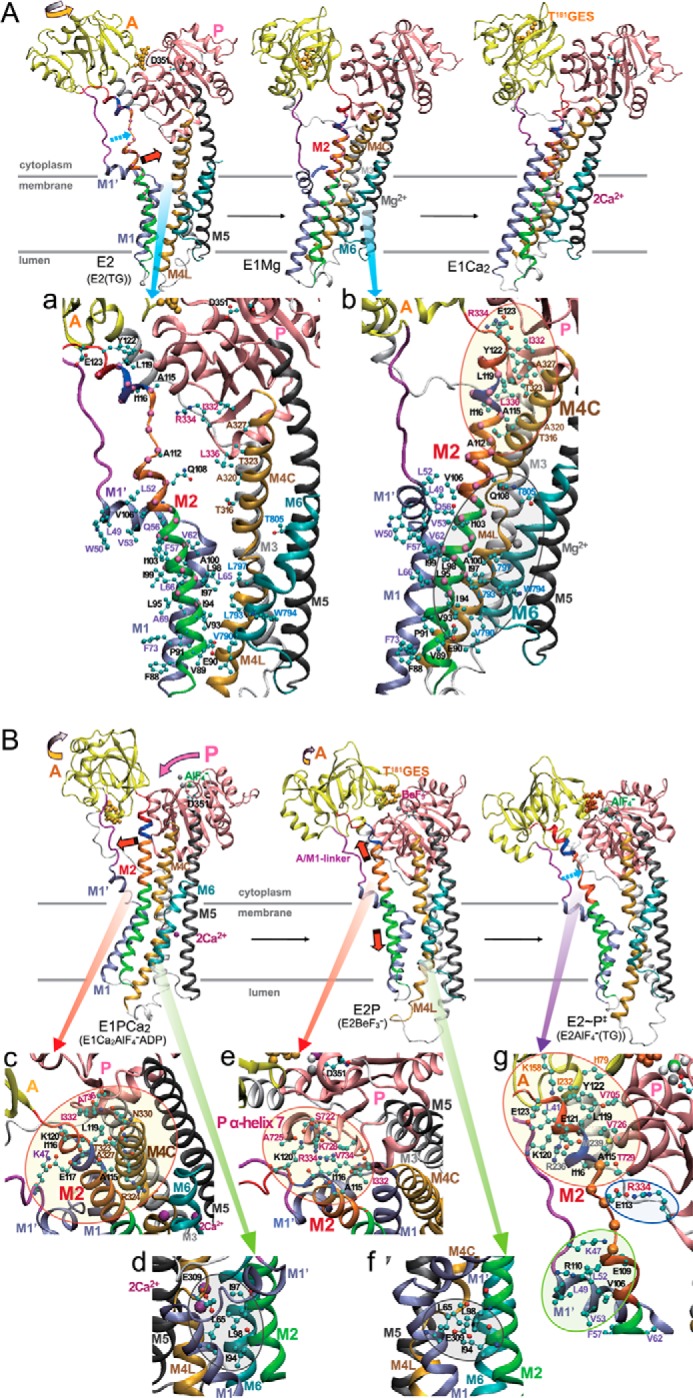
The actuator (A) domain of sarco(endo)plasmic reticulum Ca(2+)-ATPase not only plays a catalytic role but also undergoes large rotational movements that influence the distant transport sites through connections with transmembrane helices M1 and M2. Here we explore the importance of long helix M2 and its junction with the A domain by disrupting the helix structure and elongating with insertions of five glycine residues. Insertions into the membrane region of M2 and the top junctional segment impair Ca(2+) transport despite reasonable ATPase activity, indicating that they are uncoupled. These mutants fail to occlude Ca(2+). Those at the top segment also exhibited accelerated phosphoenzyme isomerization E1P → E2P. Insertions into the middle of M2 markedly accelerate E2P hydrolysis and cause strong resistance to inhibition by luminal Ca(2+). Insertions along almost the entire M2 region inhibit the dephosphorylated enzyme transition E2 → E1. The results pinpoint which parts of M2 control cytoplasm gating and which are critical for luminal gating at each stage in the transport cycle and suggest that proper gate function requires appropriate interactions, tension, and/or rigidity in the M2 region at appropriate times for coupling with A domain movements and catalysis.
Keywords: Calcium ATPase; Domain Motion; Enzyme Kinetics; Enzyme Mutation; Enzyme Structure; Phosphoenzyme Intermediate; Phosphoryl Transfer; Sarco(endo)plasmic Reticulum; Transmembrane Helix.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Glycine 105 as Pivot for a Critical Knee-like Joint between Cytoplasmic and Transmembrane Segments of the Second Transmembrane Helix in Ca2+-ATPase.J Biol Chem. 2016 Nov 18;291(47):24688-24701. doi: 10.1074/jbc.M116.759704. Epub 2016 Oct 12. J Biol Chem. 2016. PMID: 27733680 Free PMC article.
-
Critical role of Glu40-Ser48 loop linking actuator domain and first transmembrane helix of Ca2+-ATPase in Ca2+ deocclusion and release from ADP-insensitive phosphoenzyme.J Biol Chem. 2007 Nov 23;282(47):34429-47. doi: 10.1074/jbc.M707665200. Epub 2007 Sep 19. J Biol Chem. 2007. PMID: 17881350
-
Deletions of any single residues in Glu40-Ser48 loop connecting a domain and the first transmembrane helix of sarcoplasmic reticulum Ca(2+)-ATPase result in almost complete inhibition of conformational transition and hydrolysis of phosphoenzyme intermediate.J Biol Chem. 2003 Oct 3;278(40):39197-204. doi: 10.1074/jbc.M305200200. Epub 2003 Jul 11. J Biol Chem. 2003. PMID: 12857730
-
[Mechanism of ca(2+) pump as revealed by mutations, development of stable analogs of phosphorylated intermediates, and their structural analyses].Yakugaku Zasshi. 2010 Feb;130(2):179-89. doi: 10.1248/yakushi.130.179. Yakugaku Zasshi. 2010. PMID: 20118641 Review. Japanese.
-
Ca2+ -ATPase structure in the E1 and E2 conformations: mechanism, helix-helix and helix-lipid interactions.Biochim Biophys Acta. 2002 Oct 11;1565(2):246-66. doi: 10.1016/s0005-2736(02)00573-4. Biochim Biophys Acta. 2002. PMID: 12409199 Review.
Cited by
-
The N Terminus of Sarcolipin Plays an Important Role in Uncoupling Sarco-endoplasmic Reticulum Ca2+-ATPase (SERCA) ATP Hydrolysis from Ca2+ Transport.J Biol Chem. 2015 May 29;290(22):14057-67. doi: 10.1074/jbc.M115.636738. Epub 2015 Apr 16. J Biol Chem. 2015. PMID: 25882845 Free PMC article.
-
Glycine 105 as Pivot for a Critical Knee-like Joint between Cytoplasmic and Transmembrane Segments of the Second Transmembrane Helix in Ca2+-ATPase.J Biol Chem. 2016 Nov 18;291(47):24688-24701. doi: 10.1074/jbc.M116.759704. Epub 2016 Oct 12. J Biol Chem. 2016. PMID: 27733680 Free PMC article.
-
Angle change of the A-domain in a single SERCA1a molecule detected by defocused orientation imaging.Sci Rep. 2021 Jul 1;11(1):13672. doi: 10.1038/s41598-021-92986-3. Sci Rep. 2021. PMID: 34211016 Free PMC article.
-
Sarcolipin Promotes Uncoupling of the SERCA Ca2+ Pump by Inducing a Structural Rearrangement in the Energy-Transduction Domain.Biochemistry. 2016 Nov 8;55(44):6083-6086. doi: 10.1021/acs.biochem.6b00728. Epub 2016 Oct 28. Biochemistry. 2016. PMID: 27731980 Free PMC article.
-
Membrane Perturbation of ADP-insensitive Phosphoenzyme of Ca2+-ATPase Modifies Gathering of Transmembrane Helix M2 with Cytoplasmic Domains and Luminal Gating.Sci Rep. 2017 Jan 24;7:41172. doi: 10.1038/srep41172. Sci Rep. 2017. PMID: 28117348 Free PMC article.
References
-
- Toyoshima C. (2008) Structural aspects of ion pumping by Ca2+-ATPase of sarcoplasmic reticulum. Arch. Biochem. Biophys. 476, 3–11 - PubMed
-
- Toyoshima C. (2009) How Ca2+-ATPase pumps ions across the sarcoplasmic reticulum membrane. Biochim. Biophys. Acta 1793, 941–946 - PubMed
-
- Møller J. V., Olesen C., Winther A.-M. L., Nissen P. (2010) The sarcoplasmic Ca2+-ATPase: design of a perfect chemi-osmotic pump. Q. Rev. Biophys. 43, 501–566 - PubMed
-
- Toyoshima C., Nakasako M., Nomura H., Ogawa H. (2000) Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 Å resolution. Nature 405, 647–655 - PubMed
-
- Danko S., Yamasaki K., Daiho T., Suzuki H., Toyoshima C. (2001) Organization of cytoplasmic domains of sarcoplasmic reticulum Ca2+-ATPase in E1P and E1ATP states: a limited proteolysis study. FEBS Lett. 505, 129–135 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
