

A unified design space of synthetic stripe-forming networks
- PMID: 25247316
- PMCID: PMC4172969
- DOI: 10.1038/ncomms5905
A unified design space of synthetic stripe-forming networks
Abstract
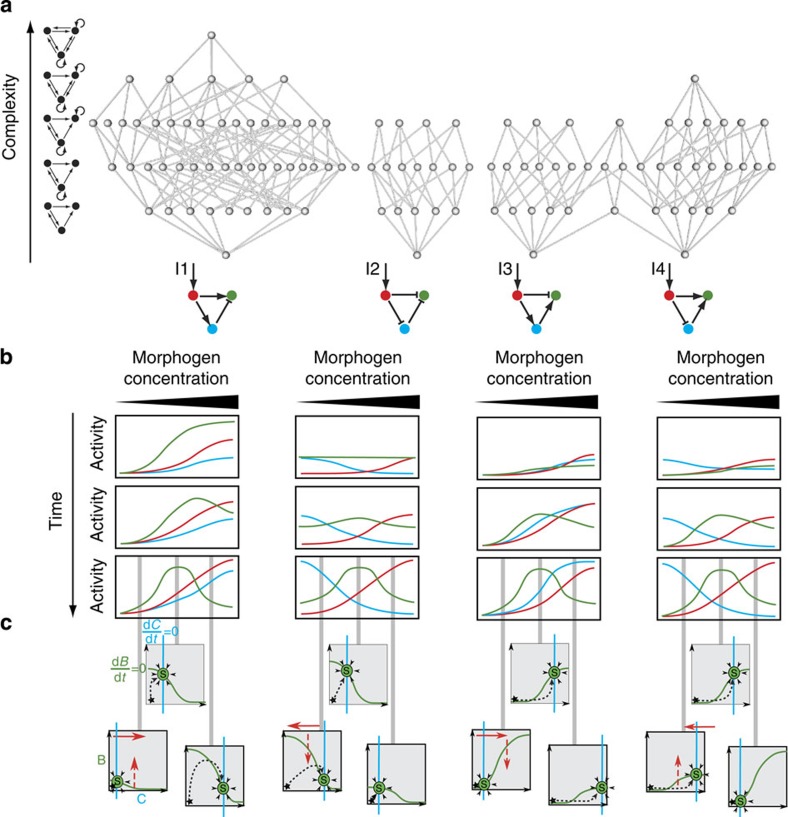
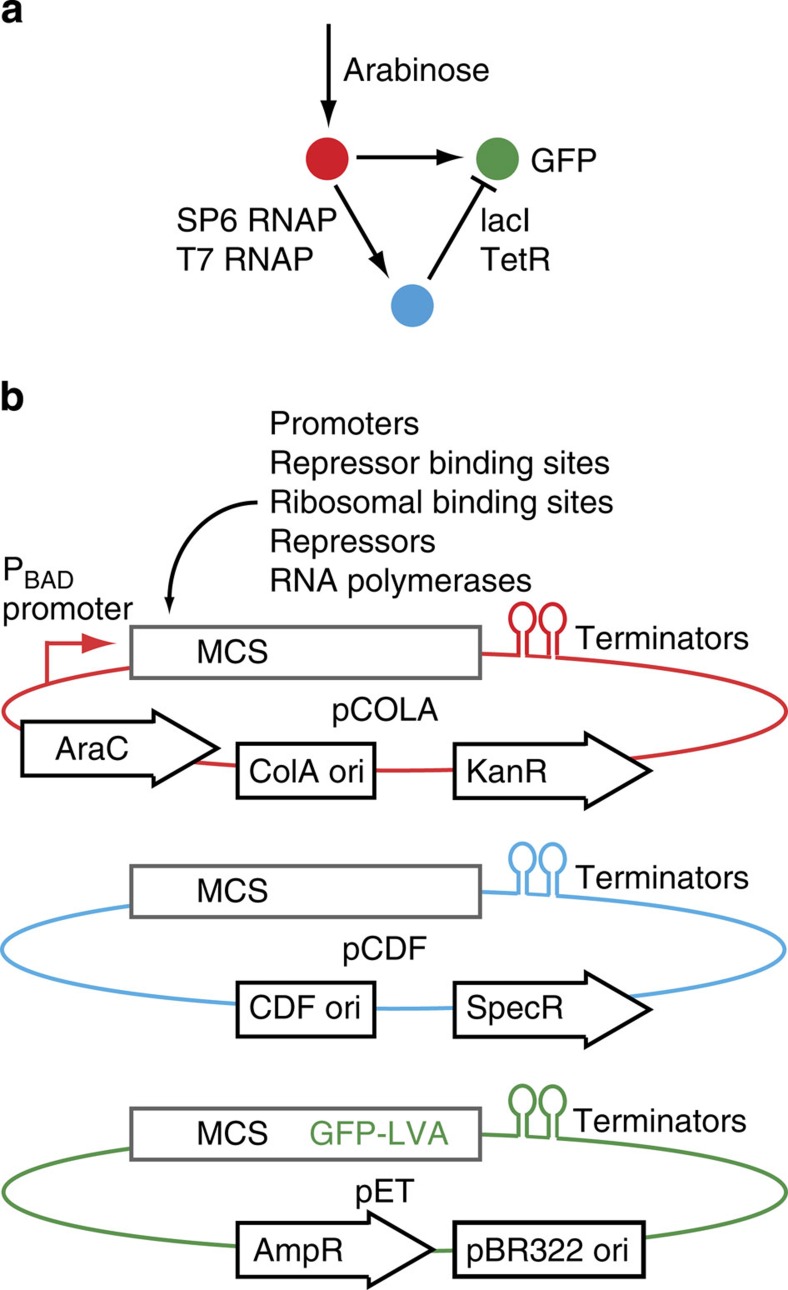
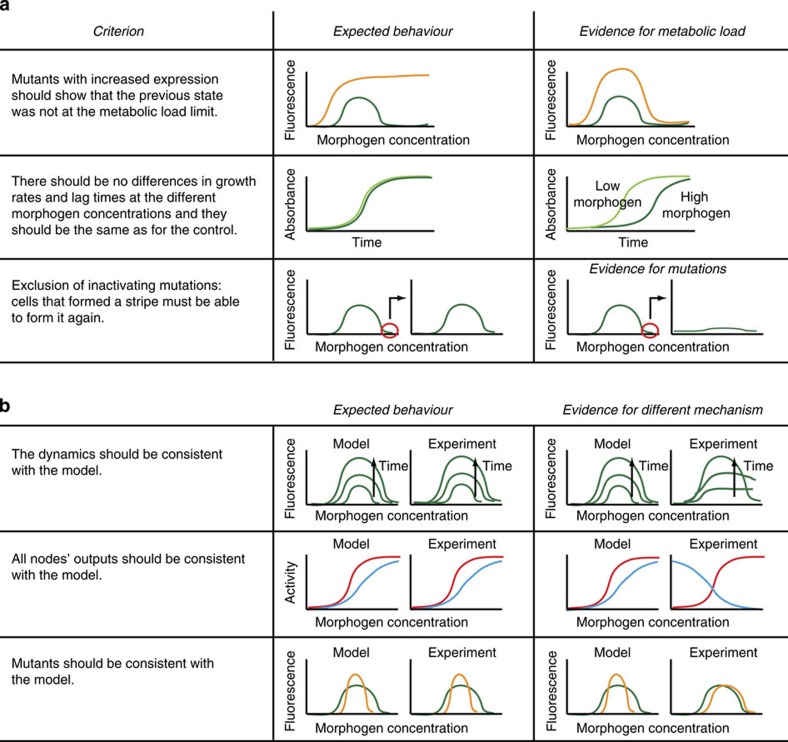
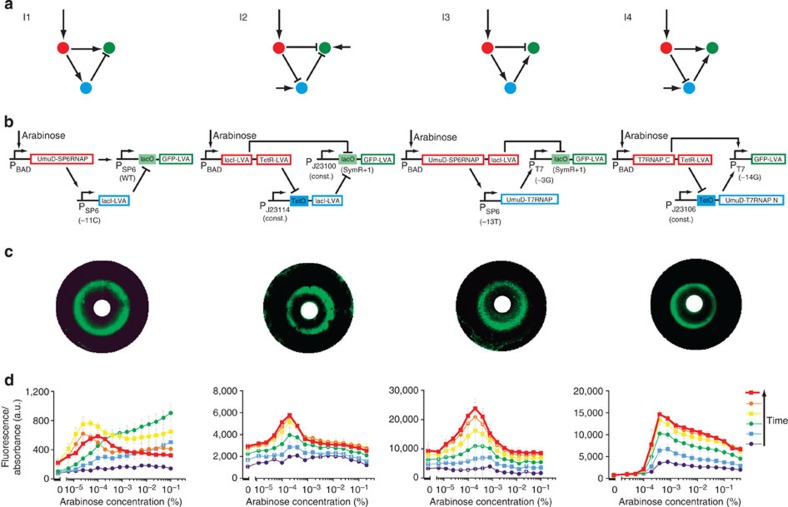
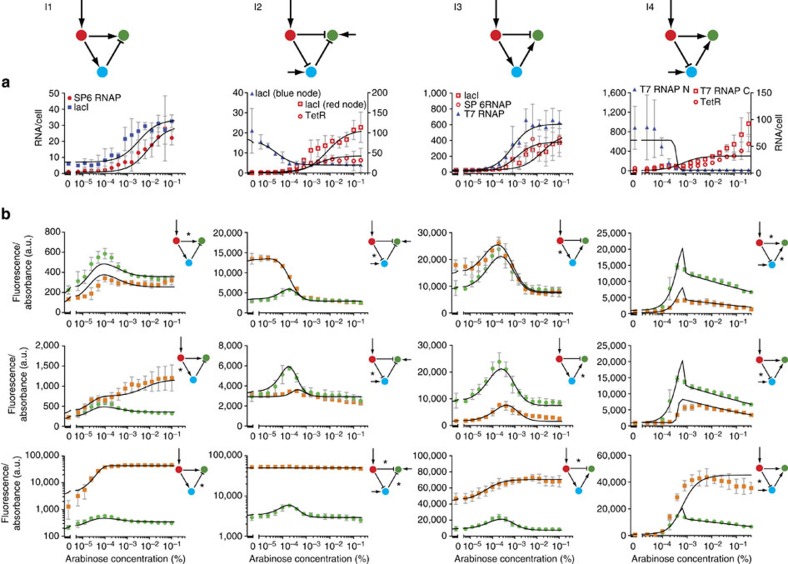
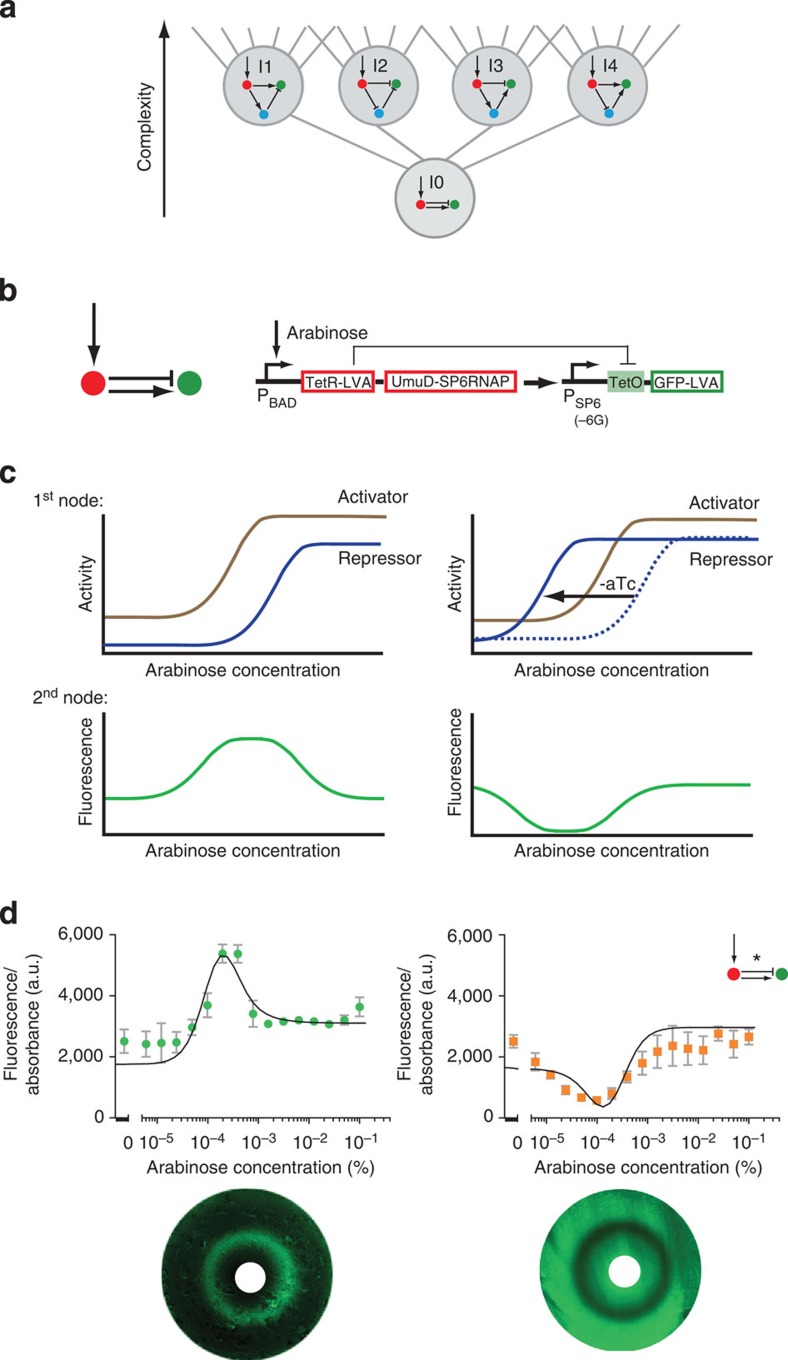
Synthetic biology is a promising tool to study the function and properties of gene regulatory networks. Gene circuits with predefined behaviours have been successfully built and modelled, but largely on a case-by-case basis. Here we go beyond individual networks and explore both computationally and synthetically the design space of possible dynamical mechanisms for 3-node stripe-forming networks. First, we computationally test every possible 3-node network for stripe formation in a morphogen gradient. We discover four different dynamical mechanisms to form a stripe and identify the minimal network of each group. Next, with the help of newly established engineering criteria we build these four networks synthetically and show that they indeed operate with four fundamentally distinct mechanisms. Finally, this close match between theory and experiment allows us to infer and subsequently build a 2-node network that represents the archetype of the explored design space.
Figures
Similar articles
-
A Framework for the Modular and Combinatorial Assembly of Synthetic Gene Circuits.ACS Synth Biol. 2019 Jul 19;8(7):1691-1697. doi: 10.1021/acssynbio.9b00174. Epub 2019 Jun 24. ACS Synth Biol. 2019. PMID: 31185158
-
Insulated transcriptional elements enable precise design of genetic circuits.Nat Commun. 2017 Jul 3;8(1):52. doi: 10.1038/s41467-017-00063-z. Nat Commun. 2017. PMID: 28674389 Free PMC article.
-
Construction of synthetic gene circuits in the Escherichia coli genome.Methods Mol Biol. 2013;1073:157-68. doi: 10.1007/978-1-62703-625-2_13. Methods Mol Biol. 2013. PMID: 23996446
-
Synthetic gene networks in mammalian cells.Curr Opin Biotechnol. 2010 Oct;21(5):690-6. doi: 10.1016/j.copbio.2010.07.006. Epub 2010 Aug 4. Curr Opin Biotechnol. 2010. PMID: 20691580 Review.
-
Engineering Synthetic Gene Circuits in Living Cells with CRISPR Technology.Trends Biotechnol. 2016 Jul;34(7):535-547. doi: 10.1016/j.tibtech.2015.12.014. Epub 2016 Jan 22. Trends Biotechnol. 2016. PMID: 26809780 Review.
Cited by
-
Collective Space-Sensing Coordinates Pattern Scaling in Engineered Bacteria.Cell. 2016 Apr 21;165(3):620-30. doi: 10.1016/j.cell.2016.03.006. Cell. 2016. PMID: 27104979 Free PMC article.
-
Synthetic cell-based materials extract positional information from morphogen gradients.Sci Adv. 2022 Apr 8;8(14):eabl9228. doi: 10.1126/sciadv.abl9228. Epub 2022 Apr 8. Sci Adv. 2022. PMID: 35394842 Free PMC article.
-
Rational engineering of synthetic microbial systems: from single cells to consortia.Curr Opin Microbiol. 2018 Oct;45:92-99. doi: 10.1016/j.mib.2018.02.009. Epub 2018 Mar 22. Curr Opin Microbiol. 2018. PMID: 29574330 Free PMC article. Review.
-
Automatic design of gene regulatory mechanisms for spatial pattern formation.NPJ Syst Biol Appl. 2024 Apr 2;10(1):35. doi: 10.1038/s41540-024-00361-5. NPJ Syst Biol Appl. 2024. PMID: 38565850 Free PMC article.
-
Engineering orthogonal dual transcription factors for multi-input synthetic promoters.Nat Commun. 2016 Dec 16;7:13858. doi: 10.1038/ncomms13858. Nat Commun. 2016. PMID: 27982027 Free PMC article.
References
-
- Smolke C. D. Building outside of the box: iGEM and the BioBricks Foundation. Nat. Biotechnol. 27, 1099–1102 (2009). - PubMed
-
- Mutalik V. K. et al. Precise and reliable gene expression via standard transcription and translation initiation elements. Nat. Methods 10, 354–360 (2013). - PubMed
-
- Mutalik V. K. et al. Quantitative estimation of activity and quality for collections of functional genetic elements. Nat. Methods 10, 347–353 (2013). - PubMed
-
- Qi L., Haurwitz R. E., Shao W., Doudna J. A. & Arkin A. P. RNA processing enables predictable programming of gene expression. Nat. Biotechnol. 30, 1002–1006 (2012). - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
