

Functional Specialization and Flexibility in Human Association Cortex
- PMID: 25249407
- PMCID: PMC4598819
- DOI: 10.1093/cercor/bhu217
Functional Specialization and Flexibility in Human Association Cortex
Erratum in
-
Functional Specialization and Flexibility in Human Association Cortex.Cereb Cortex. 2016 Jan;26(1):465. doi: 10.1093/cercor/bhv260. Epub 2015 Oct 27. Cereb Cortex. 2016. PMID: 26508334 Free PMC article. No abstract available.
Abstract
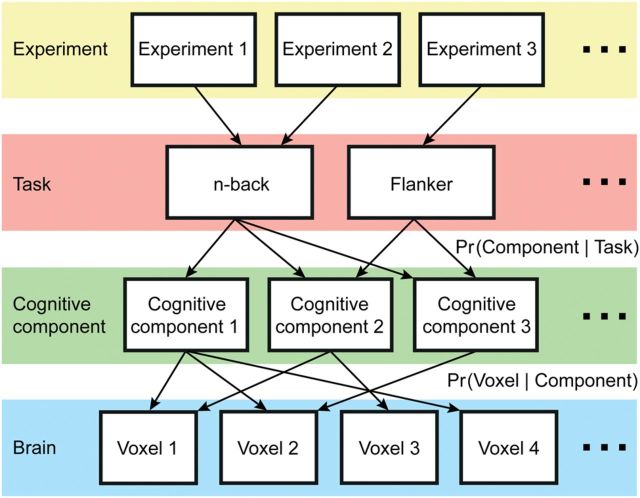
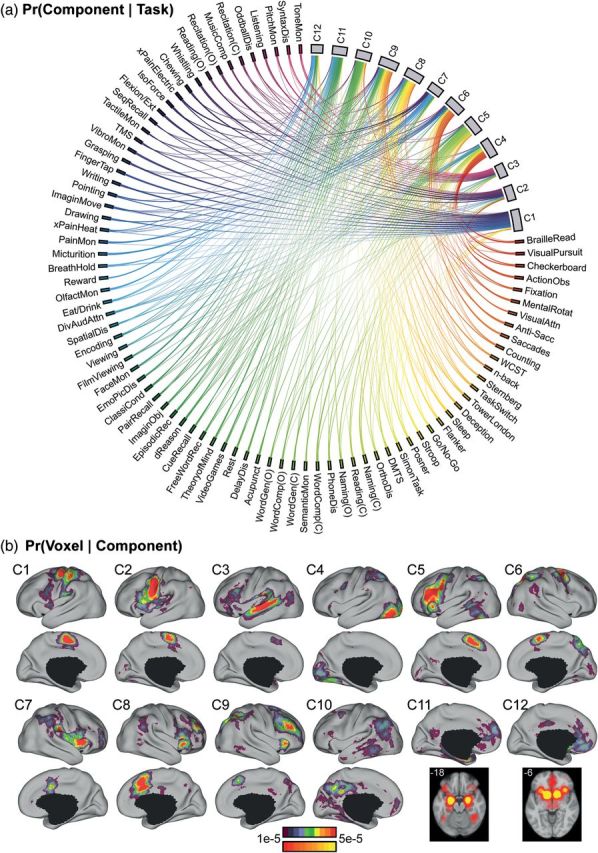
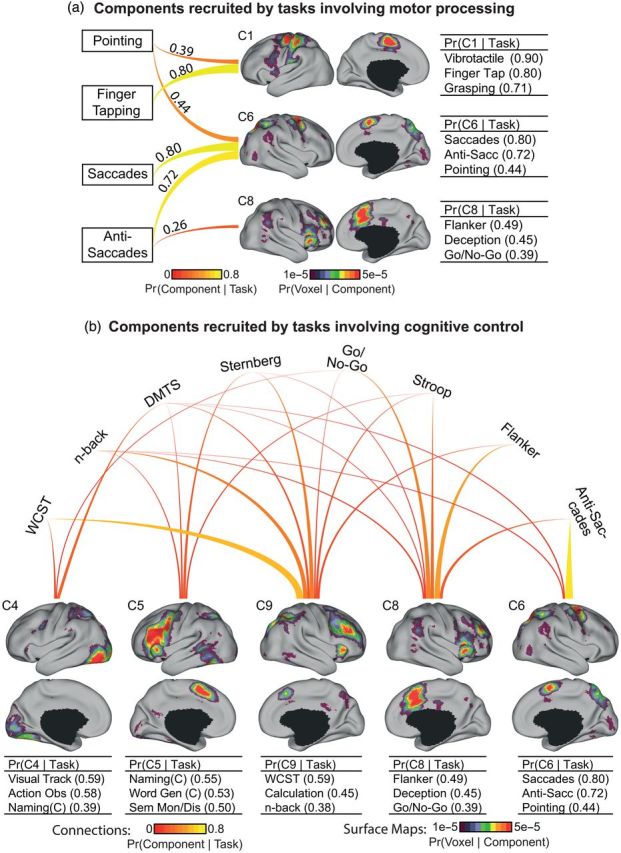
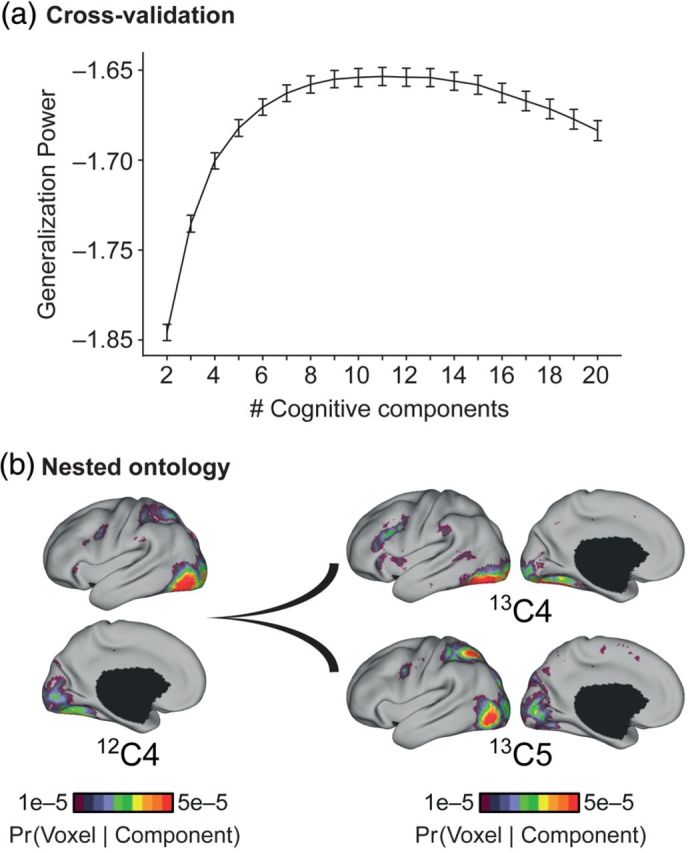
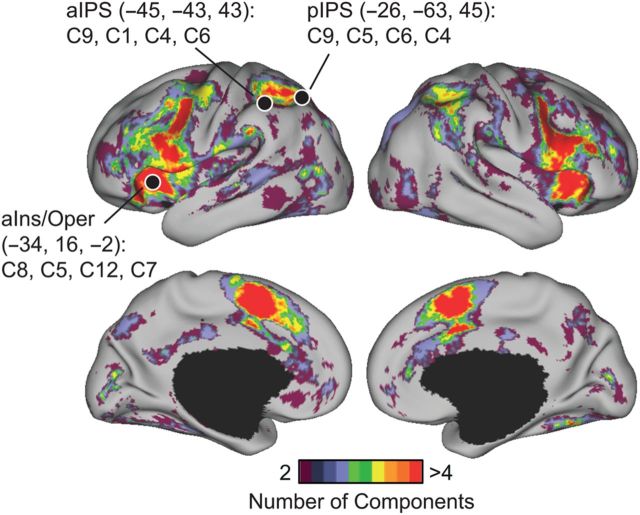
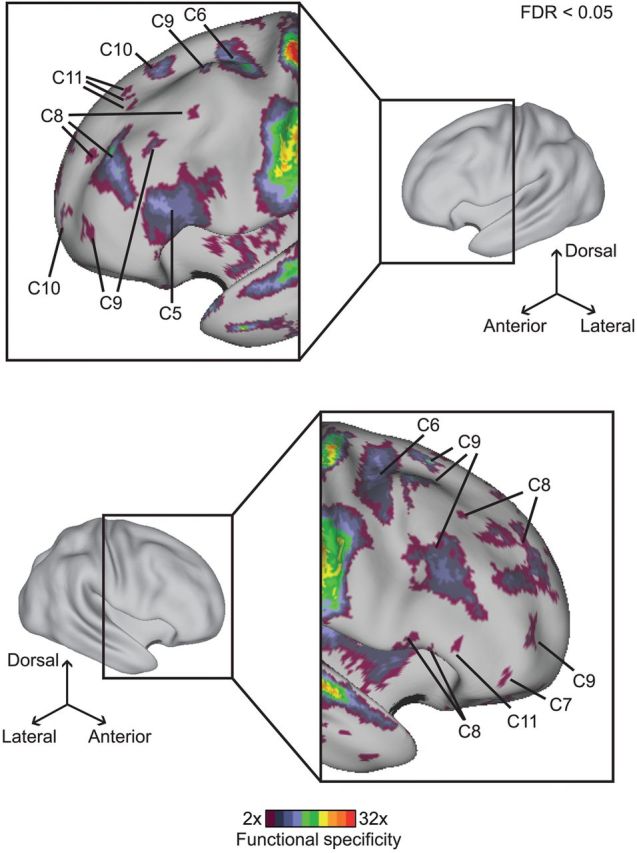
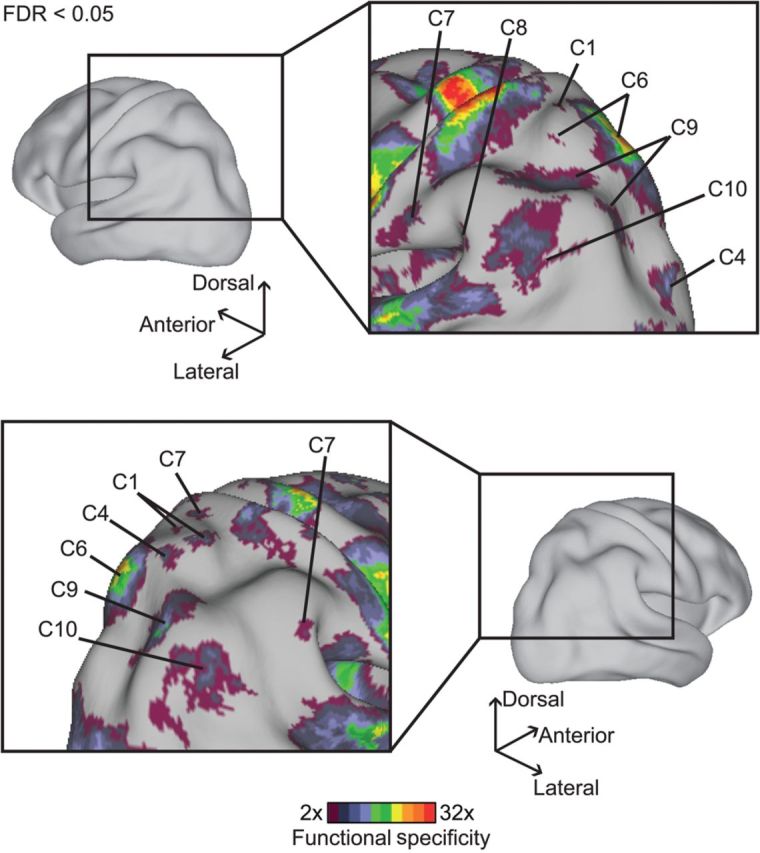
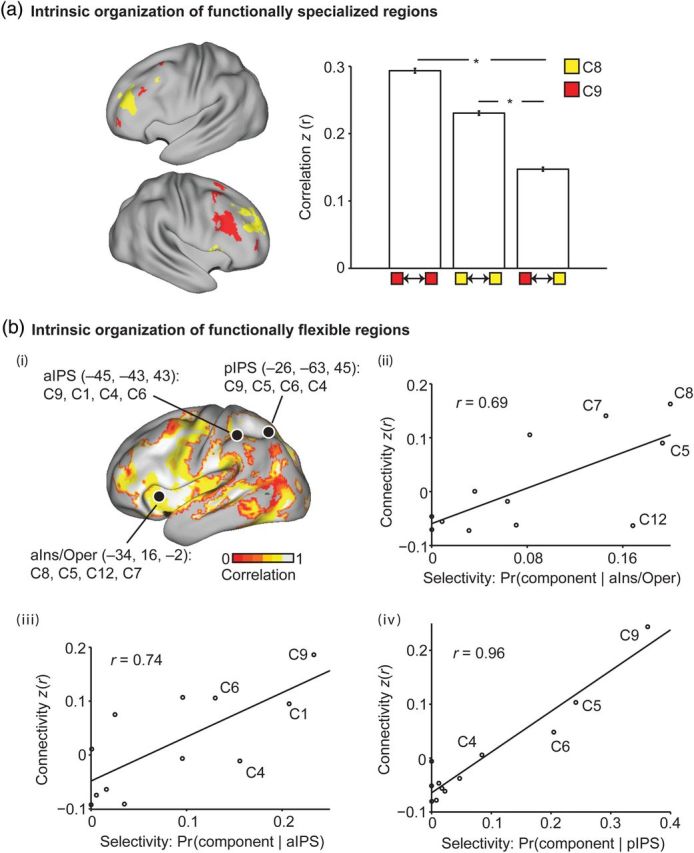
The association cortex supports cognitive functions enabling flexible behavior. Here, we explored the organization of human association cortex by mathematically formalizing the notion that a behavioral task engages multiple cognitive components, which are in turn supported by multiple overlapping brain regions. Application of the model to a large data set of neuroimaging experiments (N = 10 449) identified complex zones of frontal and parietal regions that ranged from being highly specialized to highly flexible. The network organization of the specialized and flexible regions was explored with an independent resting-state fMRI data set (N = 1000). Cortical regions specialized for the same components were strongly coupled, suggesting that components function as partially isolated networks. Functionally flexible regions participated in multiple components to different degrees. This heterogeneous selectivity was predicted by the connectivity between flexible and specialized regions. Functionally flexible regions might support binding or integrating specialized brain networks that, in turn, contribute to the ability to execute multiple and varied tasks.
Keywords: cognitive ontology; functional connectivity; meta-analysis; parietal cortex; prefrontal cortex.
© The Author 2014. Published by Oxford University Press. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures

References
-
- Aron AR, Robbins TW, Poldrack RA. 2004. Inhibition and the right inferior frontal cortex. Trends Cogn Sci. 8:170–177. - PubMed
-
- Badre D, Poldrack RA, Paré-Blagoev EJ, Insler RZ, Wagner AD. 2005. Dissociable controlled retrieval and generalized selection mechanisms in ventrolateral prefrontal cortex. Neuron. 47:907–918. - PubMed
-
- Bechtel W. 2002. Decomposing the mind-brain: a long-term pursuit. Brain Mind. 3:229–242.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
