

Deletion of serum amyloid A3 improves high fat high sucrose diet-induced adipose tissue inflammation and hyperlipidemia in female mice
- PMID: 25251243
- PMCID: PMC4177399
- DOI: 10.1371/journal.pone.0108564
Deletion of serum amyloid A3 improves high fat high sucrose diet-induced adipose tissue inflammation and hyperlipidemia in female mice
Abstract
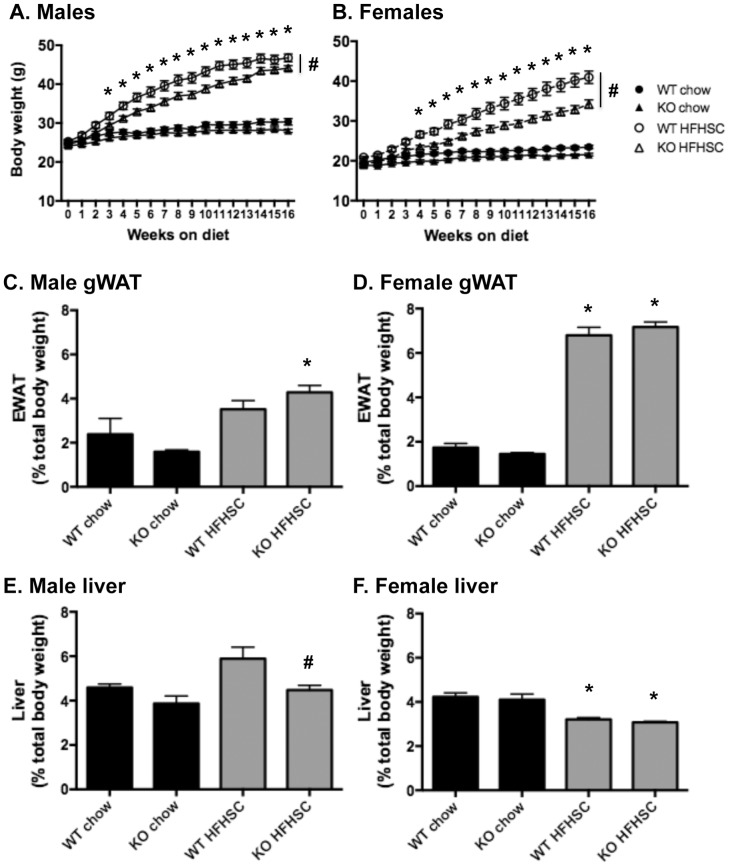
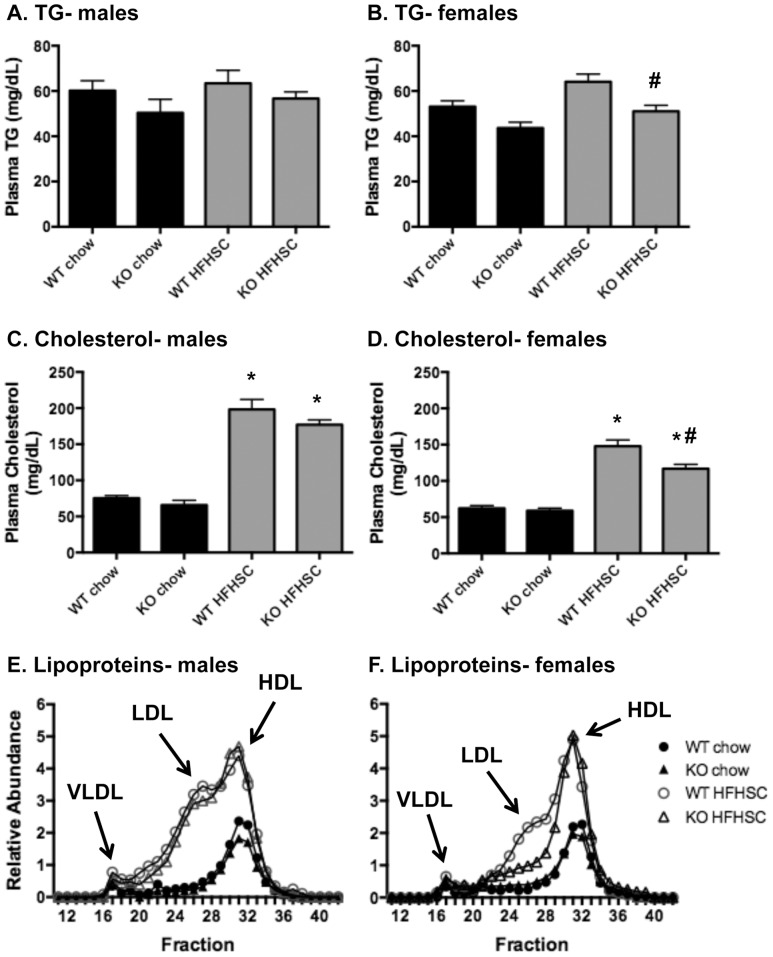
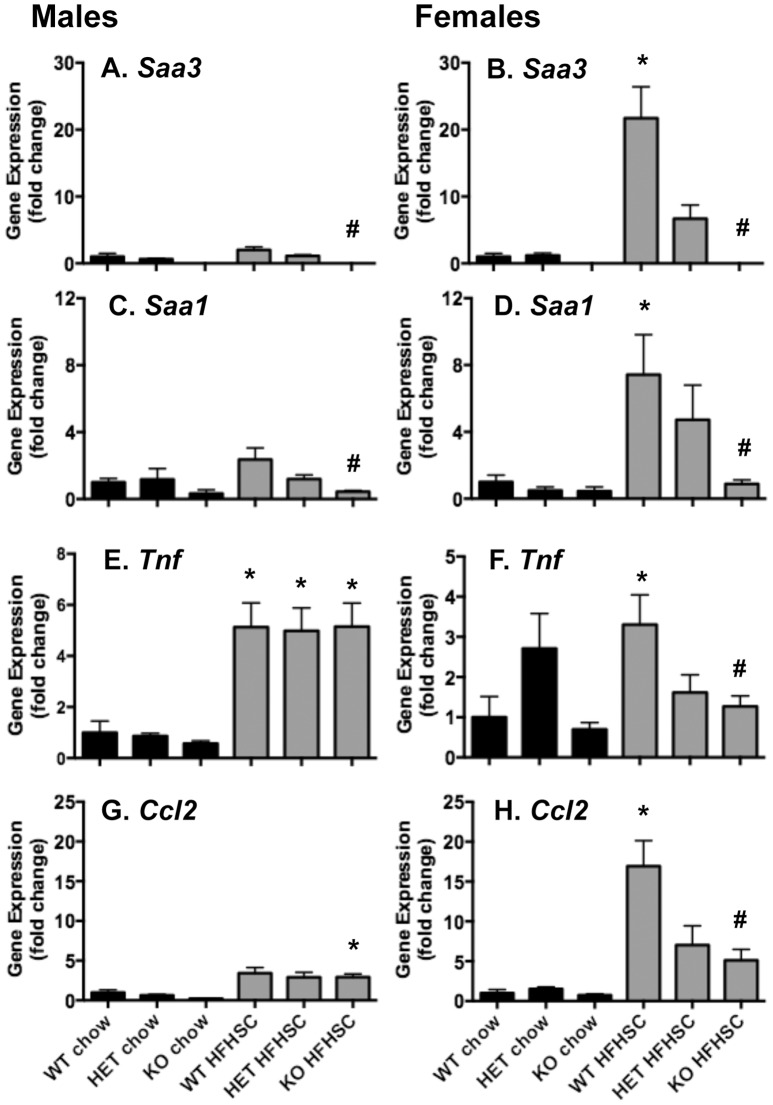
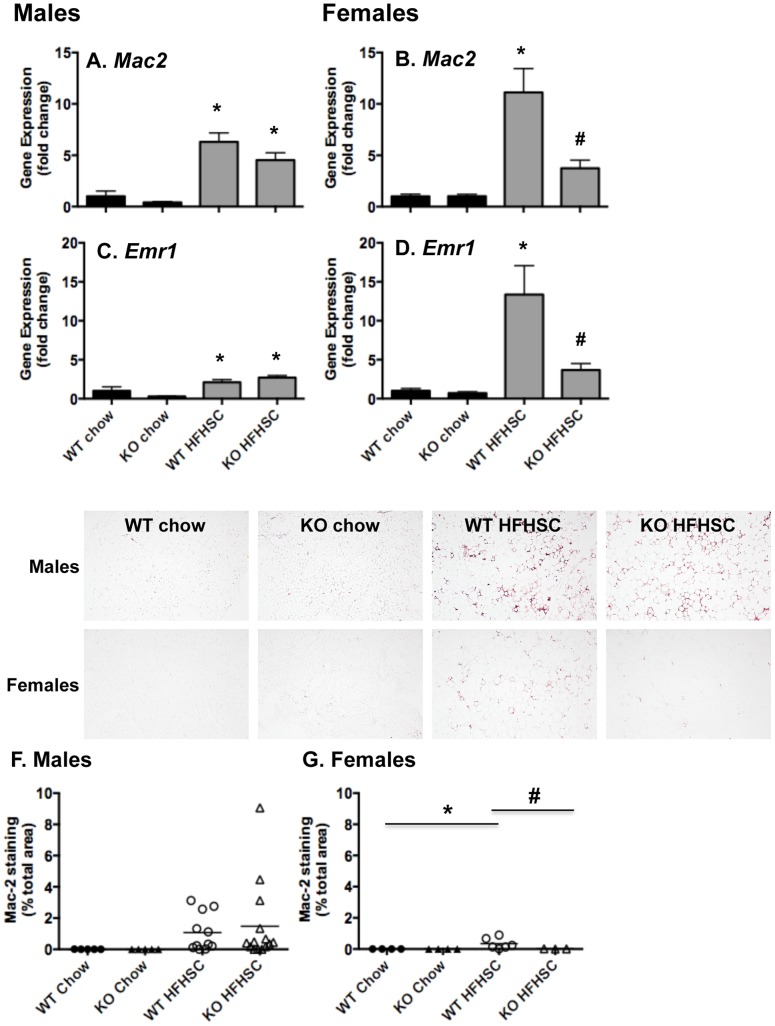
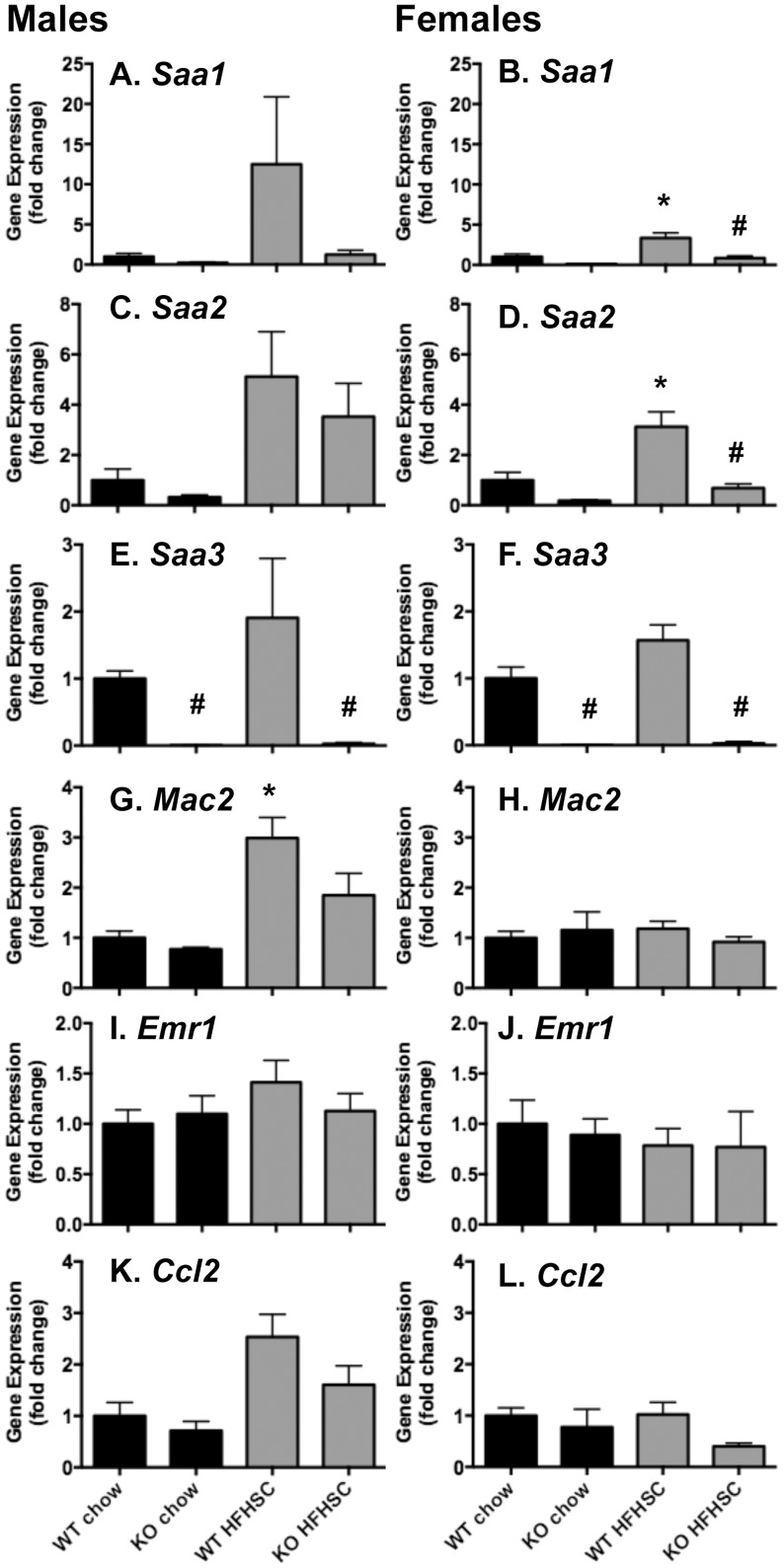
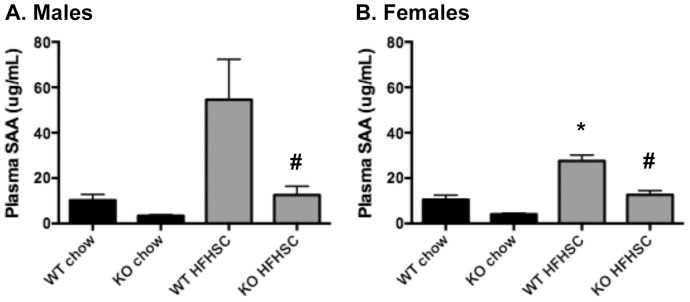
Serum amyloid A (SAA) increases in response to acute inflammatory stimuli and is modestly and chronically elevated in obesity. SAA3, an inducible form of SAA, is highly expressed in adipose tissue in obese mice where it promotes monocyte chemotaxis, providing a mechanism for the macrophage accumulation that occurs with adipose tissue expansion in obesity. Humans do not express functional SAA3 protein, but instead express SAA1 and SAA2 in hepatic as well as extrahepatic tissues, making it difficult to distinguish between liver and adipose tissue-specific SAA effects. SAA3 does not circulate in plasma, but may exert local effects that impact systemic inflammation. We tested the hypothesis that SAA3 contributes to chronic systemic inflammation and adipose tissue macrophage accumulation in obesity using mice deficient for Saa3 (Saa3(-/-)). Mice were rendered obese by feeding a pro-inflammatory high fat, high sucrose diet with added cholesterol (HFHSC). Both male and female Saa3(-/-) mice gained less weight on the HFHSC diet compared to Saa3(+/+) littermate controls, with no differences in body composition or resting metabolism. Female Saa3(-/-) mice, but not males, had reduced HFHSC diet-induced adipose tissue inflammation and macrophage content. Both male and female Saa3(-/-) mice had reduced liver Saa1 and Saa2 expression in association with reduced plasma SAA. Additionally, female Saa3(-/-) mice, but not males, showed improved plasma cholesterol, triglycerides, and lipoprotein profiles, with no changes in glucose metabolism. Taken together, these results suggest that the absence of Saa3 attenuates liver-specific SAA (i.e., SAA1/2) secretion into plasma and blunts weight gain induced by an obesogenic diet. Furthermore, adipose tissue-specific inflammation and macrophage accumulation are attenuated in female Saa3(-/-) mice, suggesting a novel sexually dimorphic role for this protein. These results also suggest that Saa3 influences liver-specific SAA1/2 expression, and that SAA3 could play a larger role in the acute phase response than previously thought.
Conflict of interest statement
Figures
References
-
- Müller S, Martin S, Koenig W, Hanifi-Moghaddam P, Rathmann W, et al. (2002) Impaired glucose tolerance is associated with increased serum concentrations of interleukin 6 and co-regulated acute-phase proteins but not TNF-alpha or its receptors. Diabetologia 45: 805–812. - PubMed
-
- Poitou C, Viguerie N, Cancello R, De Matteis R, Cinti S, et al. (2005) Serum amyloid A: production by human white adipocyte and regulation by obesity and nutrition. Diabetologia 48: 519–528. - PubMed
-
- Lewis KE, Kirk EA, McDonald TO, Wang S, Wight TN, et al. (2004) Increase in serum amyloid a evoked by dietary cholesterol is associated with increased atherosclerosis in mice. Circulation 110: 540–545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
