

Phylogenetic analysis of phenotypically characterized Cryptococcus laurentii isolates reveals high frequency of cryptic species
- PMID: 25251413
- PMCID: PMC4177401
- DOI: 10.1371/journal.pone.0108633
Phylogenetic analysis of phenotypically characterized Cryptococcus laurentii isolates reveals high frequency of cryptic species
Abstract
Background: Although Cryptococcus laurentii has been considered saprophytic and its taxonomy is still being described, several cases of human infections have already reported. This study aimed to evaluate molecular aspects of C. laurentii isolates from Brazil, Botswana, Canada, and the United States.
Methods: In this study, 100 phenotypically identified C. laurentii isolates were evaluated by sequencing the 18S nuclear ribosomal small subunit rRNA gene (18S-SSU), D1/D2 region of 28S nuclear ribosomal large subunit rRNA gene (28S-LSU), and the internal transcribed spacer (ITS) of the ribosomal region.
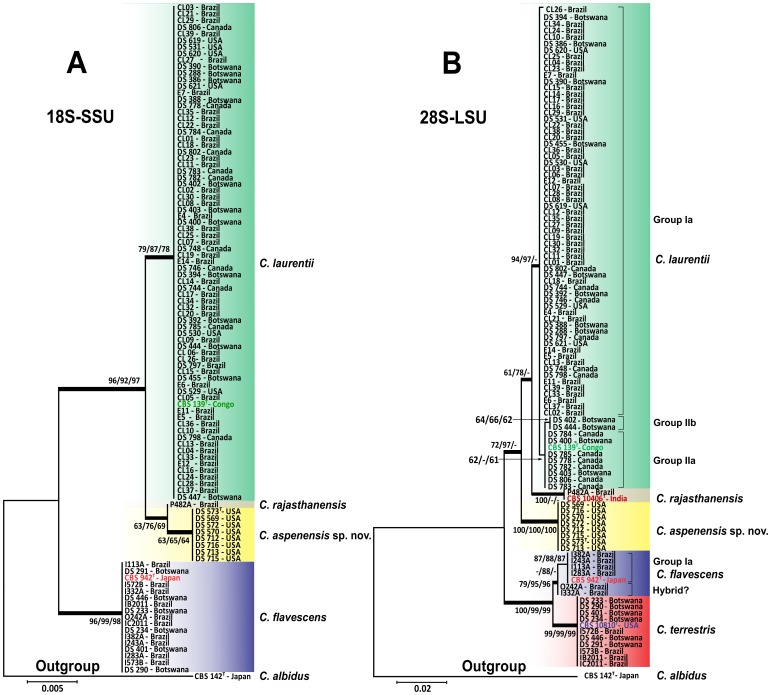
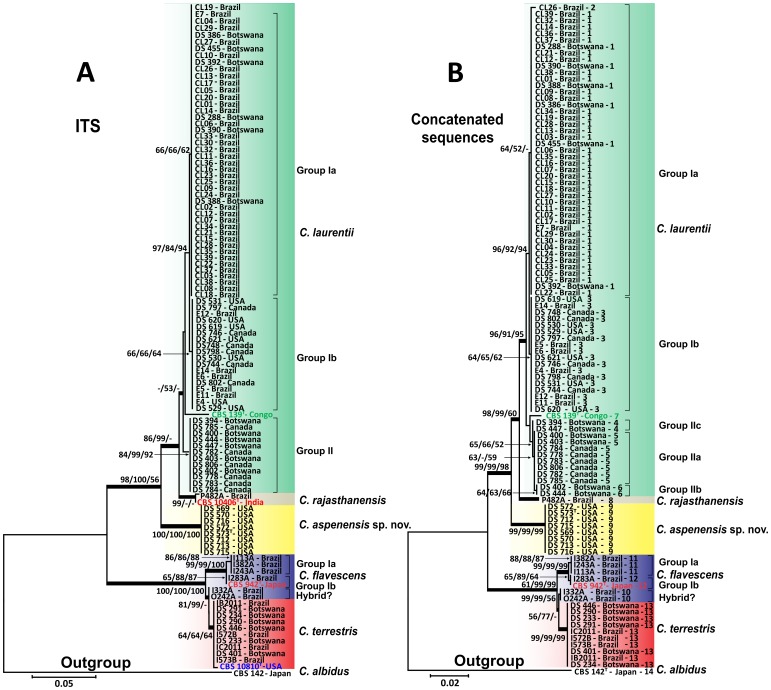
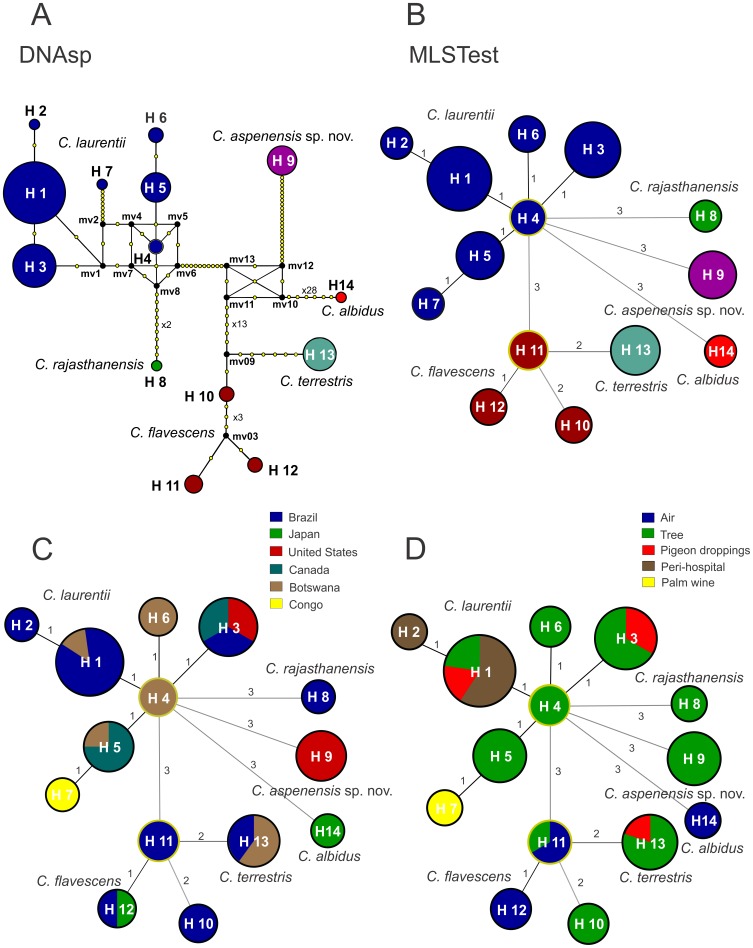
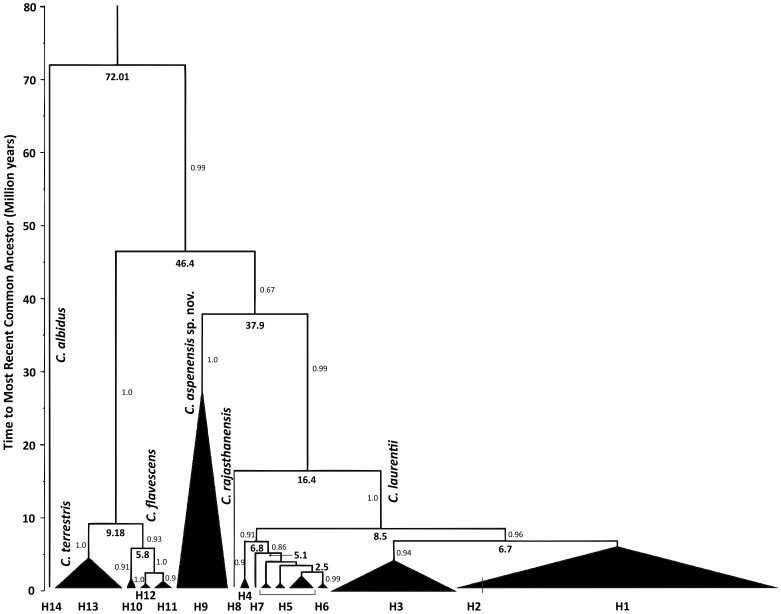
Results: BLAST searches using 550-bp, 650-bp, and 550-bp sequenced amplicons obtained from the 18S-SSU, 28S-LSU, and the ITS region led to the identification of 75 C. laurentii strains that shared 99-100% identity with C. laurentii CBS 139. A total of nine isolates shared 99% identity with both Bullera sp. VY-68 and C. laurentii RY1. One isolate shared 99% identity with Cryptococcus rajasthanensis CBS 10406, and eight isolates shared 100% identity with Cryptococcus sp. APSS 862 according to the 28S-LSU and ITS regions and designated as Cryptococcus aspenensis sp. nov. (CBS 13867). While 16 isolates shared 99% identity with Cryptococcus flavescens CBS 942 according to the 18S-SSU sequence, only six were confirmed using the 28S-LSU and ITS region sequences. The remaining 10 shared 99% identity with Cryptococcus terrestris CBS 10810, which was recently described in Brazil. Through concatenated sequence analyses, seven sequence types in C. laurentii, three in C. flavescens, one in C. terrestris, and one in the C. aspenensis sp. nov. were identified.
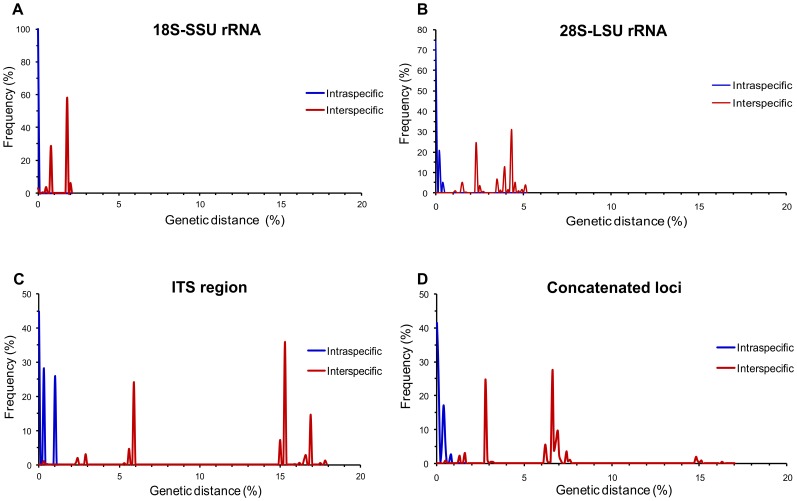
Conclusions: Sequencing permitted the characterization of 75% of the environmental C. laurentii isolates from different geographical areas and the identification of seven haplotypes of this species. Among sequenced regions, the increased variability of the ITS region in comparison to the 18S-SSU and 28S-LSU regions reinforces its applicability as a DNA barcode.
Conflict of interest statement
Figures

References
-
- Averbuch D, Boekhoutt T, Falk R, Engelhard D, Shapiro M, et al. (2002) Fungemia in a cancer patient caused by fluconazole-resistant Cryptococcus laurentii . Med Mycol 40: 479–484. - PubMed
-
- Bauters TG, Swinne D, Boekhout T, Noens L, Nelis HJ (2002) Repeated isolation of Cryptococcus laurentii from the oropharynx of an immunocompromized patient. Mycopathologia 153: 133–135. - PubMed
-
- Manfredi R, Fulgaro C, Sabbatani S, Legnani G, Fasulo G (2006) Emergence of amphotericin B-resistant Cryptococcus laurentii meningoencephalitis shortly after treatment for Cryptococcus neoformans meningitis in a patient with AIDS. AIDS Patient Care STDS 20: 227–232. - PubMed
-
- Kordossis T, Avlami A, Velegraki A, Stefanou I, Georgakopoulos G, et al. (1998) First report of Cryptococcus laurentii meningitis and a fatal case of Cryptococcus albidus cryptococcaemia in AIDS patients. Med Mycol 36: 335–339. - PubMed
-
- Banerjee P, Haider M, Trehan V, Mishra B, Thakur A, et al. (2013) Cryptococcus laurentii fungemia. Indian J Med Microbiol 31: 75–77. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
