

Phosphate concentration and arbuscular mycorrhizal colonisation influence the growth, yield and expression of twelve PHT1 family phosphate transporters in foxtail millet (Setaria italica)
- PMID: 25251671
- PMCID: PMC4177549
- DOI: 10.1371/journal.pone.0108459
Phosphate concentration and arbuscular mycorrhizal colonisation influence the growth, yield and expression of twelve PHT1 family phosphate transporters in foxtail millet (Setaria italica)
Abstract
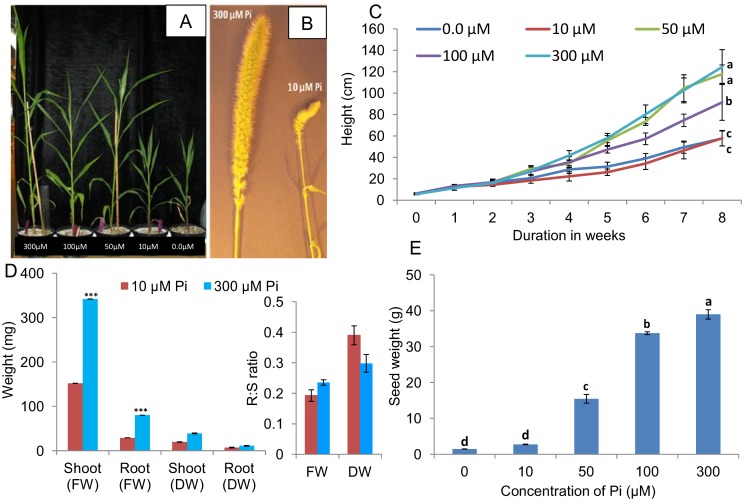
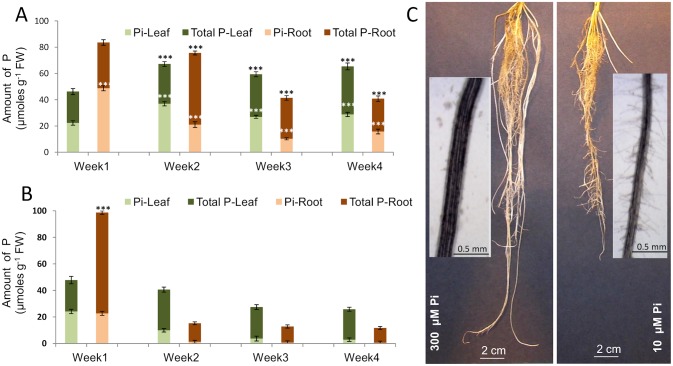
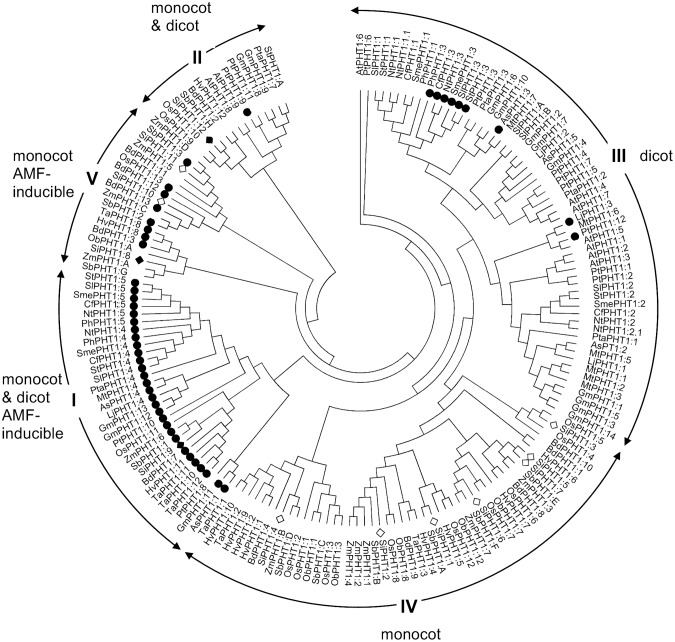
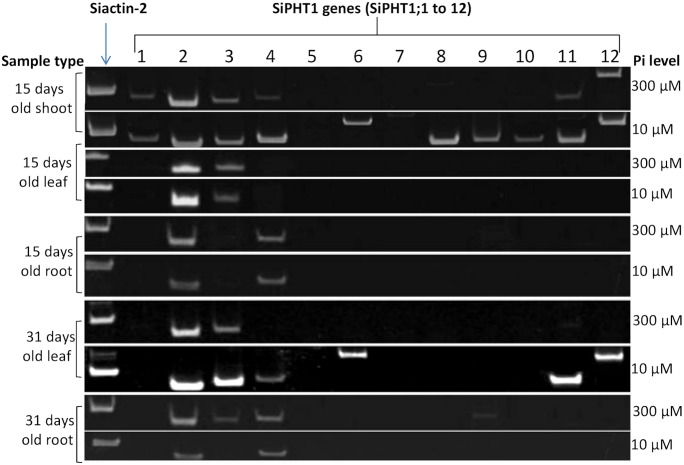
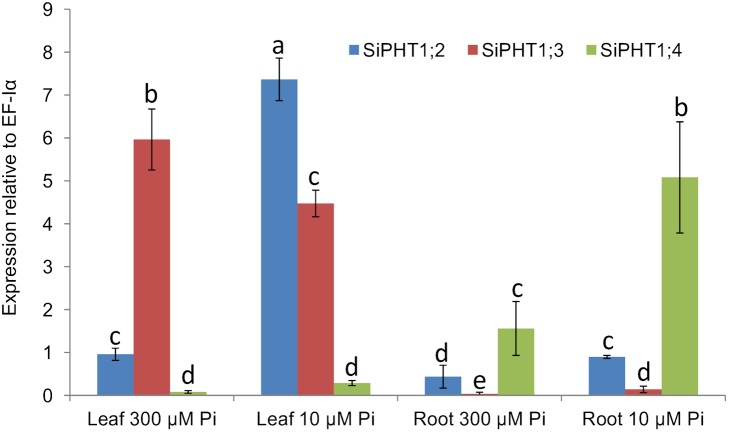
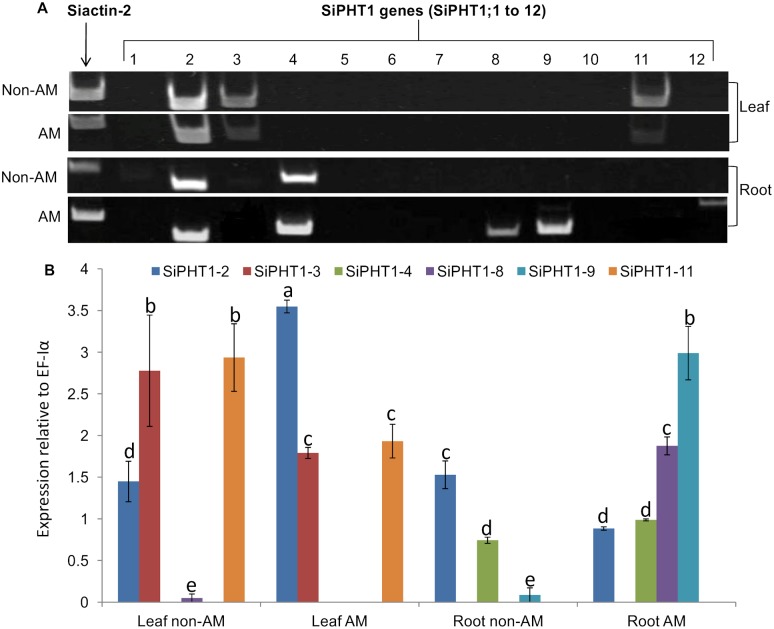
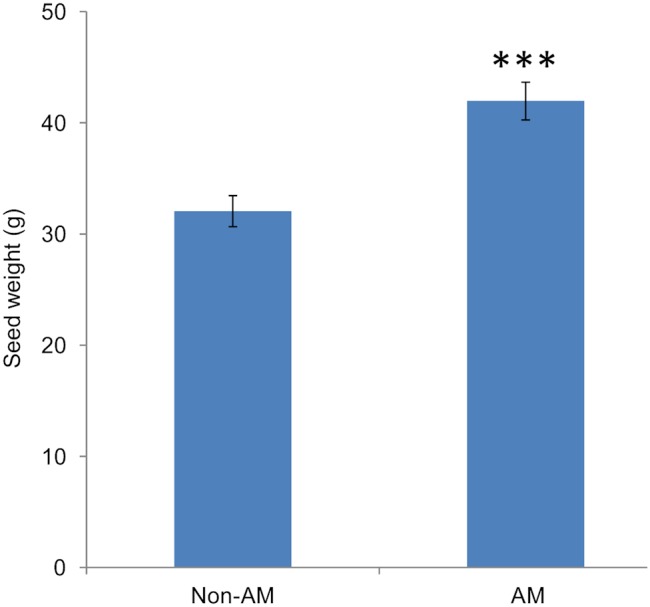
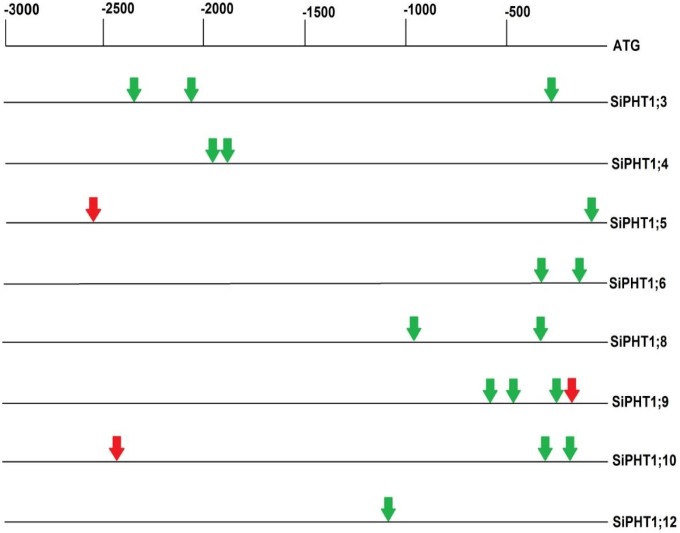
Phosphorus (P) is an essential element which plays several key roles in all living organisms. Setaria italica (foxtail millet) is a model species for panacoid grasses including several millet species widely grown in arid regions of Asia and Africa, and for the bioenergy crop switchgrass. The growth responses of S. italica to different levels of inorganic phosphate (Pi) and to colonisation with the arbuscular mycorrhizal fungus Funneliformis mosseae (syn. Glomus mosseae) were studied. Phosphate is taken up from the environment by the PHT1 family of plant phosphate transporters, which have been well characterized in several plant species. Bioinformatic analysis identified 12 members of the PHT1 gene family (SiPHT1;1-1;12) in S. italica, and RT and qPCR analysis showed that most of these transporters displayed specific expression patterns with respect to tissue, phosphate status and arbuscular mycorrhizal colonisation. SiPHT1;2 was found to be expressed in all tissues and in all growth conditions tested. In contrast, expression of SiPHT1;4 was induced in roots after 15 days growth in hydroponic medium of low Pi concentration. Expression of SiPHT1;8 and SiPHT1;9 in roots was selectively induced by colonisation with F. mosseae. SiPHT1;3 and SiPHT1;4 were found to be predominantly expressed in leaf and root tissues respectively. Several other transporters were expressed in shoots and leaves during growth in low Pi concentrations. This study will form the basis for the further characterization of these transporters, with the long term goal of improving the phosphate use efficiency of foxtail millet.
Conflict of interest statement
Figures
References
-
- Raghothama KG (1999) Phosphate acquisition. Annual Review of Plant Physiology and Plant Molecular Biology 50: 665–693. - PubMed
-
- Atkinson D (1973) Some general effects of phosphorus deficiency on growth and development. New Phytologist 72: 101–111.
-
- Sicher RC, Kremer DF (1988) Effects of phosphate deficiency on assimilate partitioning in barley seedlings. Plant Science 57: 9–17.
-
- Hajabbasi MA, Schumacher TE (1994) Phosphorus effects on root-growth and development in 2 maize genotypes. Plant and Soil 158: 39–46.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
