

A context-independent N-glycan signal targets the misfolded extracellular domain of Arabidopsis STRUBBELIG to endoplasmic-reticulum-associated degradation
- PMID: 25251695
- PMCID: PMC4255730
- DOI: 10.1042/BJ20141057
A context-independent N-glycan signal targets the misfolded extracellular domain of Arabidopsis STRUBBELIG to endoplasmic-reticulum-associated degradation
Abstract
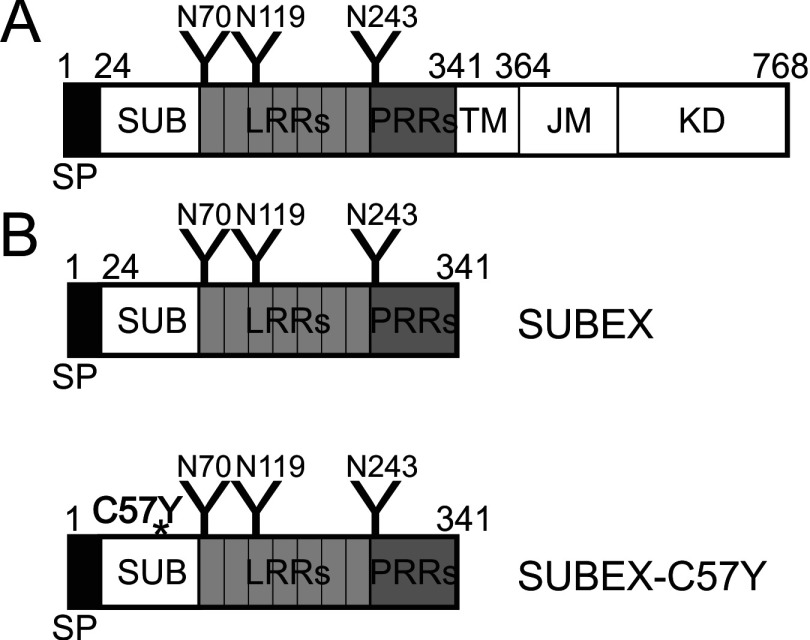
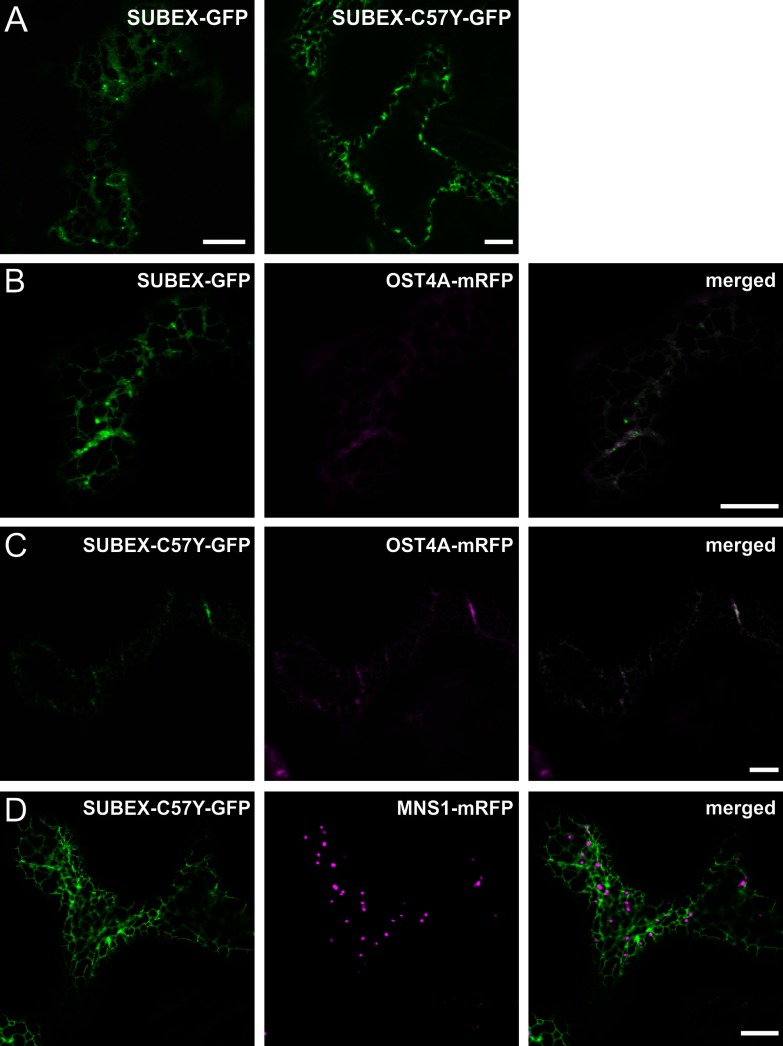
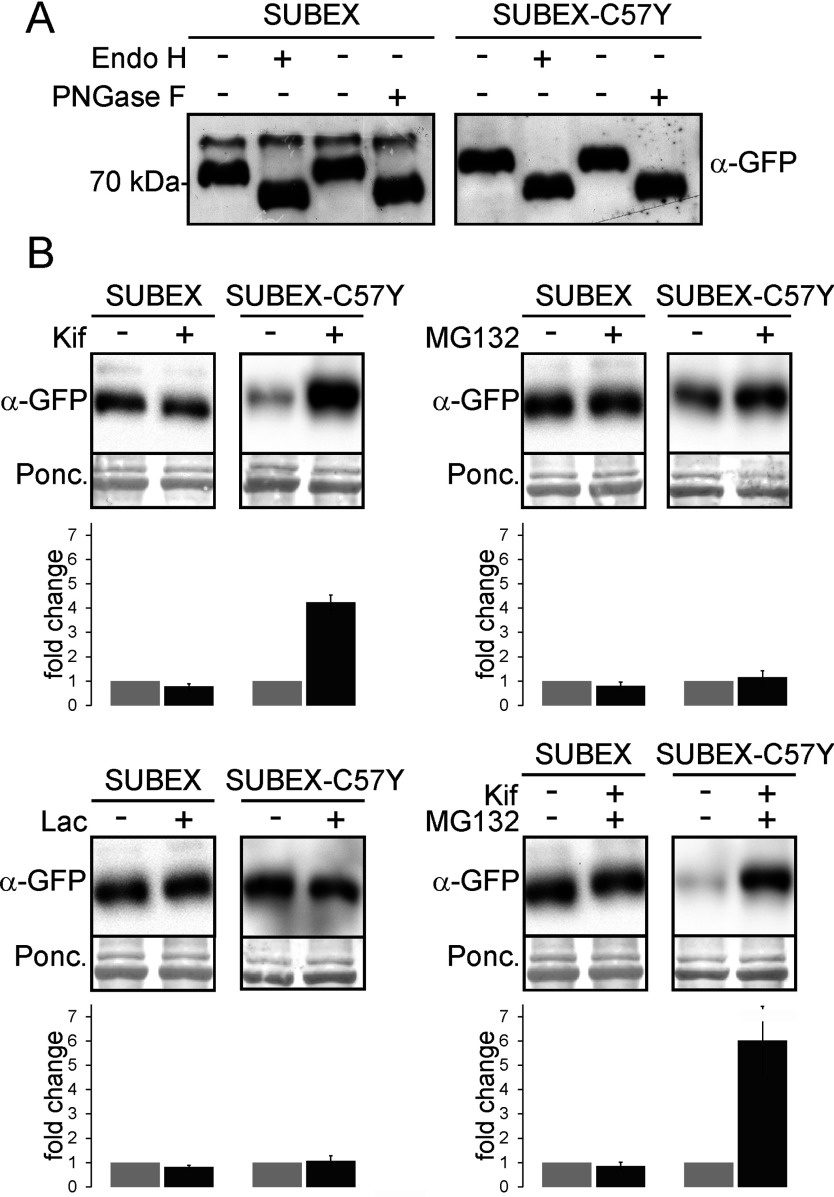
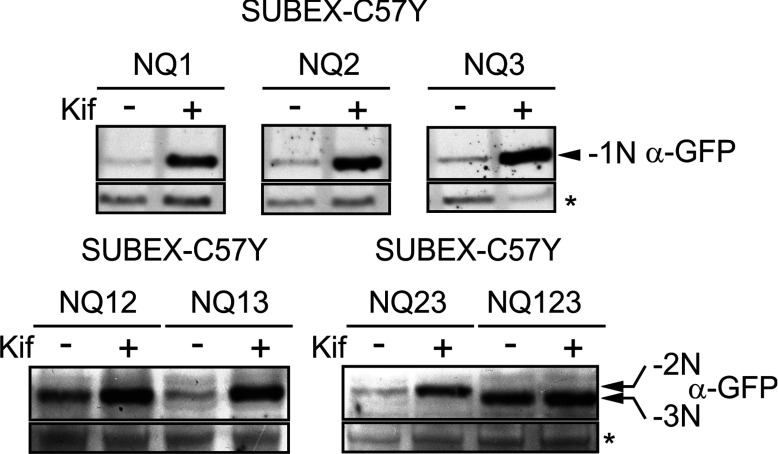
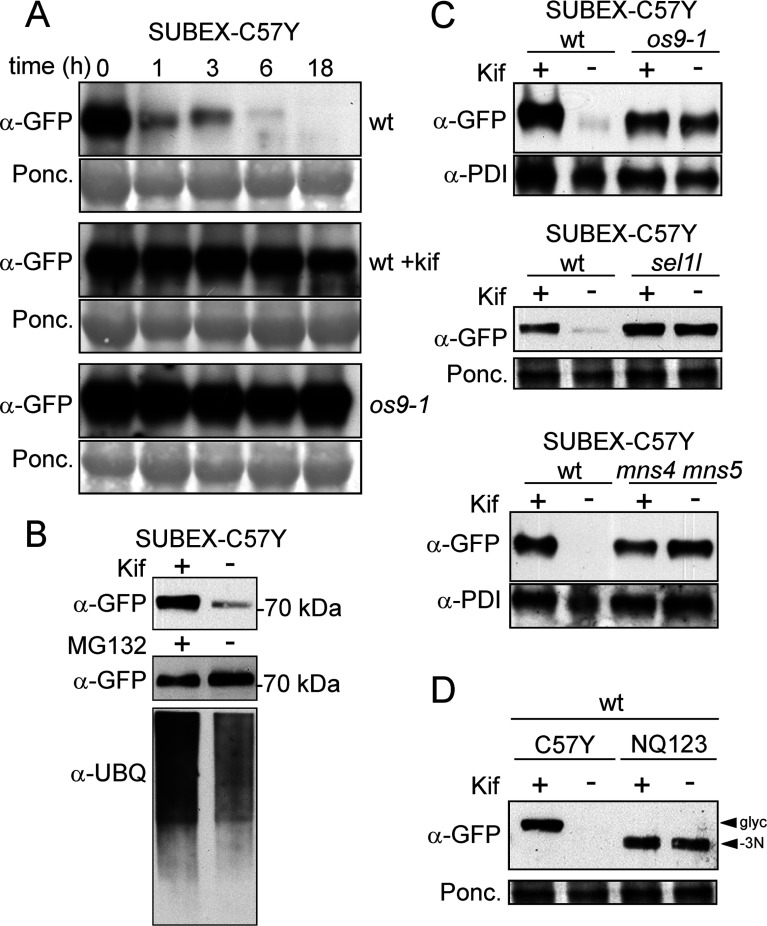
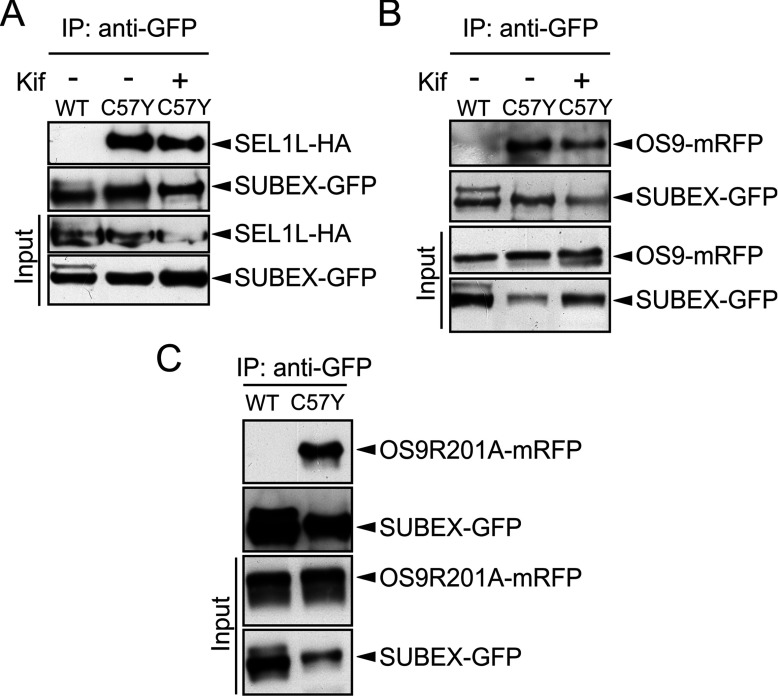
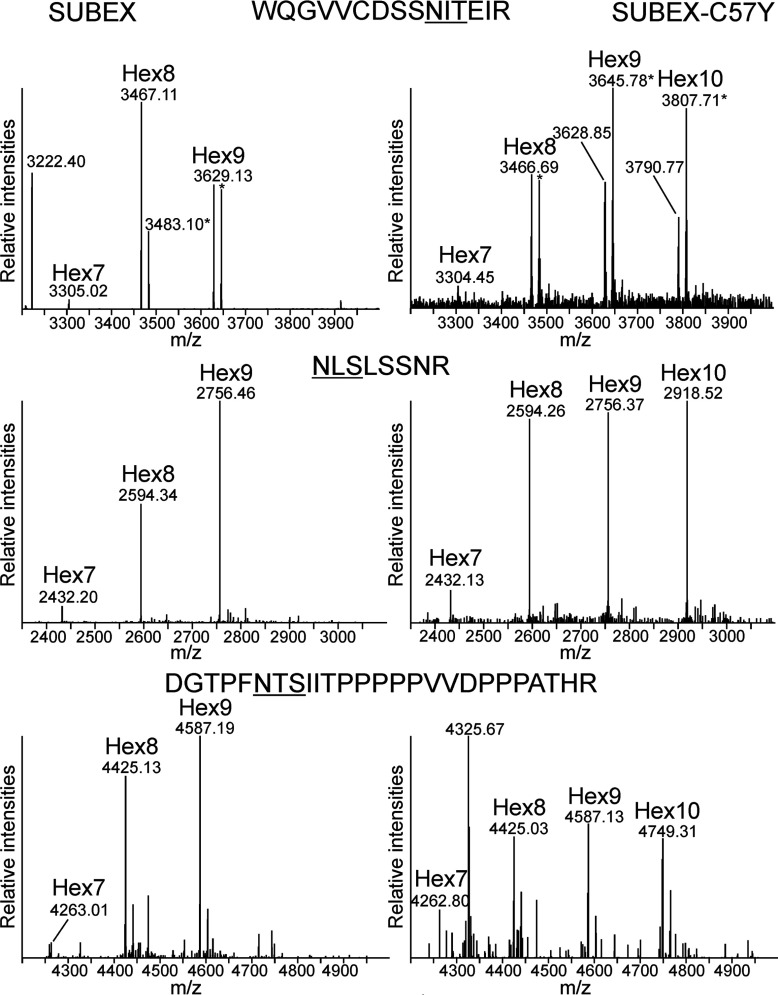
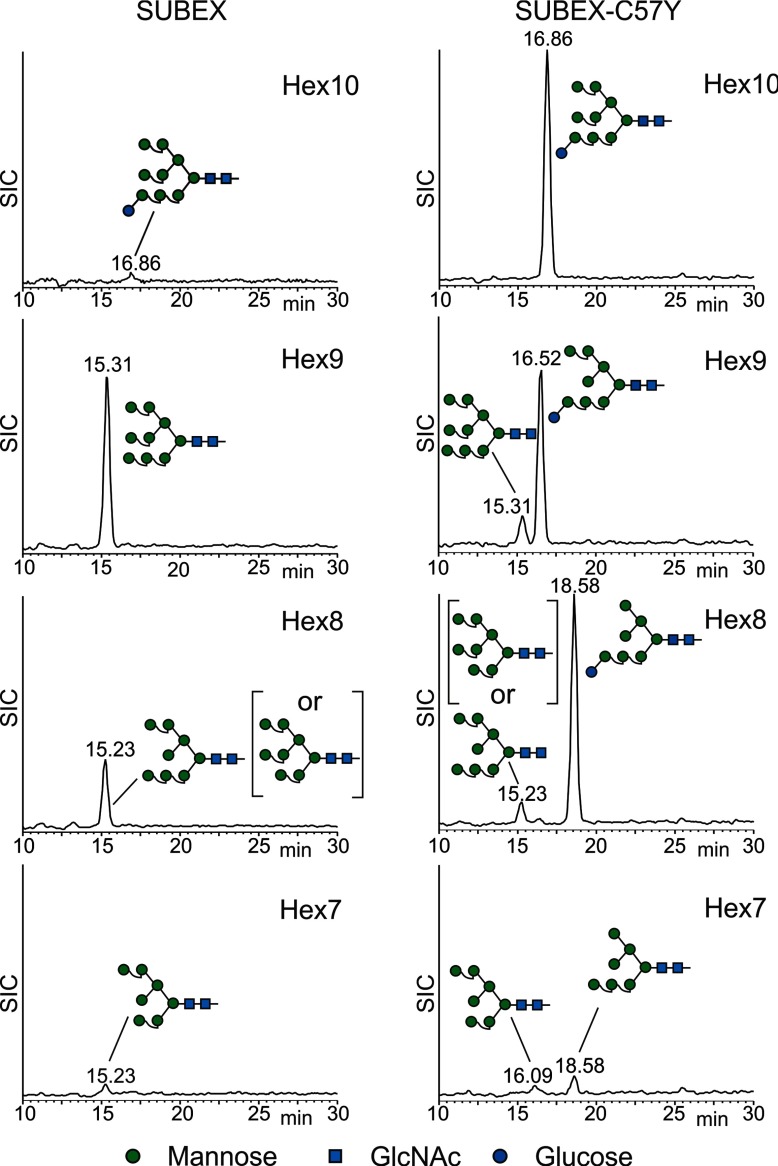
N-glycosylation of proteins plays an important role in the determination of the fate of newly synthesized glycoproteins in the endoplasmic reticulum (ER). Specific oligosaccharide structures recruit molecular chaperones that promote folding or mannose-binding lectins that assist in the clearance of improperly-folded glycoproteins by delivery to ER-associated degradation (ERAD). In plants, the mechanisms and factors that recognize non-native proteins and sort them to ERAD are poorly understood. In the present study, we provide evidence that a misfolded variant of the STRUBBELIG (SUB) extracellular domain (SUBEX-C57Y) is degraded in a glycan-dependent manner in plants. SUBEX-C57Y is an ER-retained glycoprotein with three N-glycans that is stabilized in the presence of kifunensine, a potent inhibitor of α-mannosidases. Stable expression in Arabidopsis thaliana knockout mutants revealed that SUBEX-C57Y degradation is dependent on the ER lectin OS9 and its associated ERAD factor SEL1L. SUBEX-C57Y was also stabilized in plants lacking the α-mannosidases MNS4 and MNS5 that generate a terminal α1,6-linked mannose on the C-branch of N-glycans. Notably, the glycan signal for degradation is not constrained to a specific position within SUBEX-C57Y. Structural analysis revealed that SUBEX-C57Y harbours considerable amounts of Glc1Man7GlcNAc2 N-glycans suggesting that the ER-quality control processes involving calnexin/calreticulin (CNX/CRT) and ERAD are tightly interconnected to promote protein folding or disposal by termination of futile folding attempts.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials