

A critical time window for dopamine actions on the structural plasticity of dendritic spines
- PMID: 25258080
- PMCID: PMC4225776
- DOI: 10.1126/science.1255514
A critical time window for dopamine actions on the structural plasticity of dendritic spines
Abstract
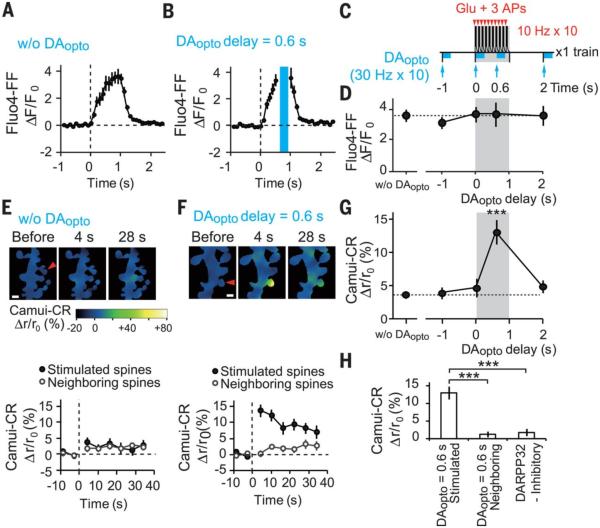
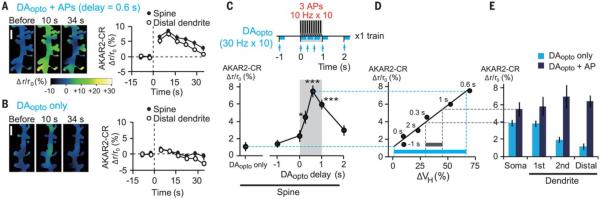
Animal behaviors are reinforced by subsequent rewards following within a narrow time window. Such reward signals are primarily coded by dopamine, which modulates the synaptic connections of medium spiny neurons in the striatum. The mechanisms of the narrow timing detection, however, remain unknown. Here, we optically stimulated dopaminergic and glutamatergic inputs separately and found that dopamine promoted spine enlargement only during a narrow time window (0.3 to 2 seconds) after the glutamatergic inputs. The temporal contingency was detected by rapid regulation of adenosine 3',5'-cyclic monophosphate in thin distal dendrites, in which protein-kinase A was activated only within the time window because of a high phosphodiesterase activity. Thus, we describe a molecular basis of reinforcement plasticity at the level of single dendritic spines.
Copyright © 2014, American Association for the Advancement of Science.
Figures


Comment in
-
Synaptic plasticity: Timing is everything.Nat Rev Neurosci. 2014 Nov;15(11):699. doi: 10.1038/nrn3850. Epub 2014 Oct 15. Nat Rev Neurosci. 2014. PMID: 25315389 No abstract available.
References
-
- Thorndike EL. Animal Intelligence. Macmillan; New York: 1911.
-
- Roitman MF, Wheeler RA, Carelli RM. Nucleus accumbens neurons are innately tuned for rewarding and aversive taste stimuli, encode their predictors, and are linked to motor output. Neuron. 2005;45:587–597. - PubMed
-
- Dan Y, Poo MM. Spike timing-dependent plasticity: From synapse to perception. Physiol. Rev. 2006;86:1033–1048. - PubMed
-
- Black J, Belluzzi JD, Stein L. Reinforcement delay of one second severely impairs acquisition of brain self-stimulation. Brain Res. 1985;359:113–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
