

A peptide derived from G0/G1 switch gene 2 acts as noncompetitive inhibitor of adipose triglyceride lipase
- PMID: 25258314
- PMCID: PMC4239610
- DOI: 10.1074/jbc.M114.602599
A peptide derived from G0/G1 switch gene 2 acts as noncompetitive inhibitor of adipose triglyceride lipase
Abstract
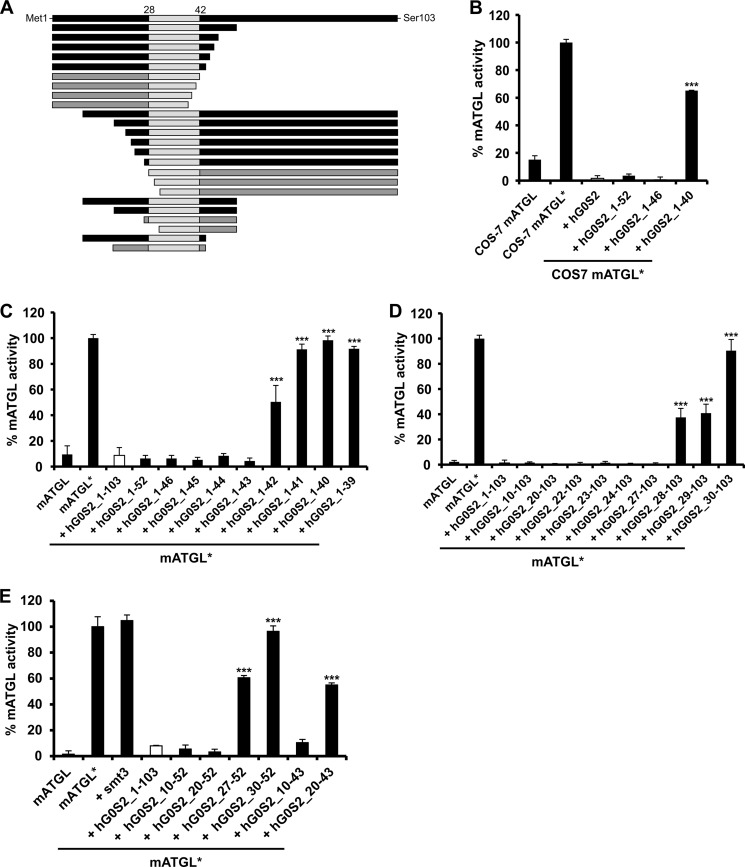
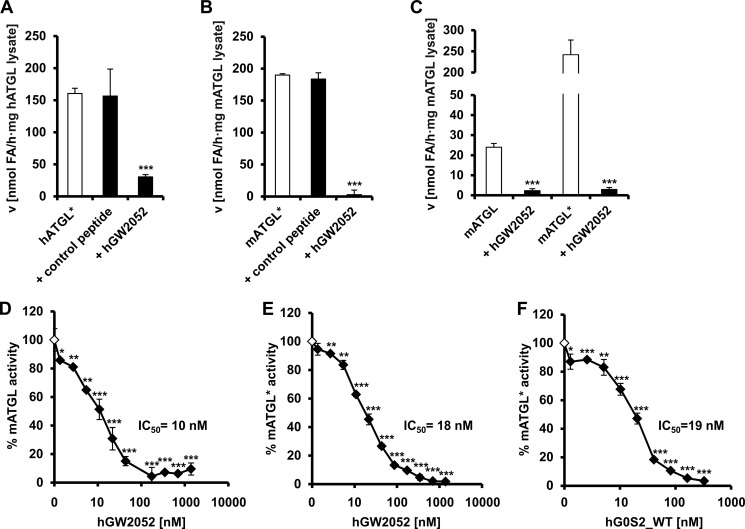
The protein G0/G1 switch gene 2 (G0S2) is a small basic protein that functions as an endogenous inhibitor of adipose triglyceride lipase (ATGL), a key enzyme in intracellular lipolysis. In this study, we identified a short sequence covering residues Lys-20 to Ala-52 in G0S2 that is still fully capable of inhibiting mouse and human ATGL. We found that a synthetic peptide corresponding to this region inhibits ATGL in a noncompetitive manner in the nanomolar range. This peptide is highly selective for ATGL and does not inhibit other lipases, including hormone-sensitive lipase, monoacylglycerol lipase, lipoprotein lipase, and patatin domain-containing phospholipases 6 and 7. Because increased lipolysis is linked to the development of metabolic disorders, the inhibition of ATGL by G0S2-derived peptides may represent a novel therapeutic tool to modulate lipolysis.
Keywords: Adipose Triglyceride Lipase (ATGL); Atglistatin; Enzyme Inhibitor; G0/G1 Switch Gene 2 (G0S2); Lipid; Lipid Metabolism; Lipolysis; Noncompetitive Inhibition; Peptides; hGW2052.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures


References
-
- Unger R. H., Clark G. O., Scherer P. E., Orci L. (2010) Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim. Biophys. Acta 1801, 209–214 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
