

μABC: a systematic microsecond molecular dynamics study of tetranucleotide sequence effects in B-DNA
- PMID: 25260586
- PMCID: PMC4231739
- DOI: 10.1093/nar/gku855
μABC: a systematic microsecond molecular dynamics study of tetranucleotide sequence effects in B-DNA
Abstract
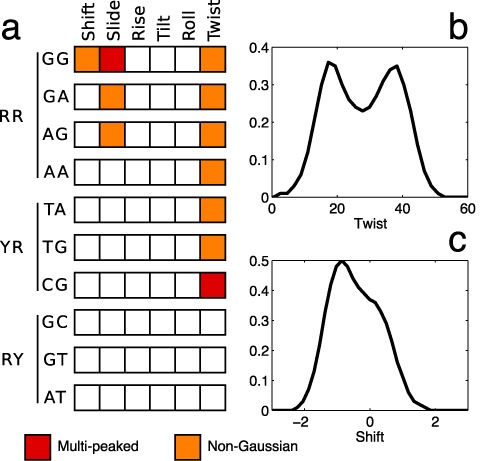
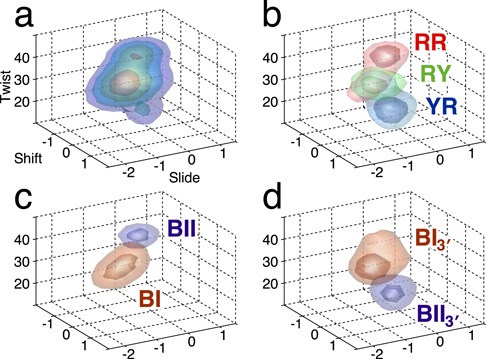
We present the results of microsecond molecular dynamics simulations carried out by the ABC group of laboratories on a set of B-DNA oligomers containing the 136 distinct tetranucleotide base sequences. We demonstrate that the resulting trajectories have extensively sampled the conformational space accessible to B-DNA at room temperature. We confirm that base sequence effects depend strongly not only on the specific base pair step, but also on the specific base pairs that flank each step. Beyond sequence effects on average helical parameters and conformational fluctuations, we also identify tetranucleotide sequences that oscillate between several distinct conformational substates. By analyzing the conformation of the phosphodiester backbones, it is possible to understand for which sequences these substates will arise, and what impact they will have on specific helical parameters.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






References
-
- Wing R., Drew H., Takano T., Broka C., Tanaka S., Itakura K., Dickerson R.E. Crystal structure analysis of a complete turn of B-DNA. Nature. 1980;287:755–758. - PubMed
-
- Juo Z.S., Chiu T.K., Leiberman P.M., Baikalov I., Berk A.J., Dickerson R.E. How proteins recognize the TATA box. J. Mol. Biol. 1996;261:239–254. - PubMed
-
- Dickerson R.E., Chiu T.K. Helix bending as a factor in protein/DNA recognition. Biopolymers. 1997;44:361–403. - PubMed
-
- Paillard G., Lavery R. Analyzing protein-DNA recognition mechanisms. Structure. 2004;12:113–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
