

High glucose dephosphorylates serine 46 and inhibits p53 apoptotic activity
- PMID: 25260780
- PMCID: PMC4181716
- DOI: 10.1186/s13046-014-0079-4
High glucose dephosphorylates serine 46 and inhibits p53 apoptotic activity
Abstract
Background: In response to diverse genotoxic stimuli p53 is activated as transcription factor to exert its tumor-suppressor function. P53 activation requires protein stabilization, nuclear localization and posttranslational modifications in key residues that may influence p53 selection of target genes. Among them, serine 46 (Ser46) phosphorylation is considered specific for apoptotic activation. Hyperglicaemia, the high blood glucose condition, may negatively affect tumor response to therapies through several mechanisms, conferring resistance to drug-induced cell death. However, whether high glucose might modify p53Ser46 phosphorylation has never been addressed.
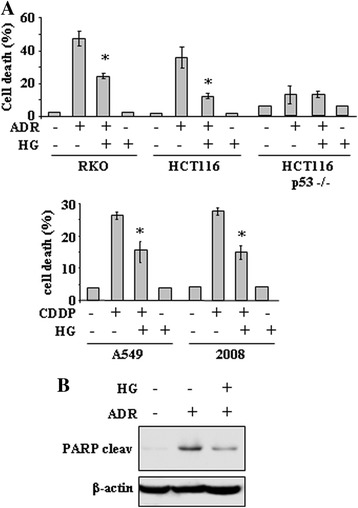
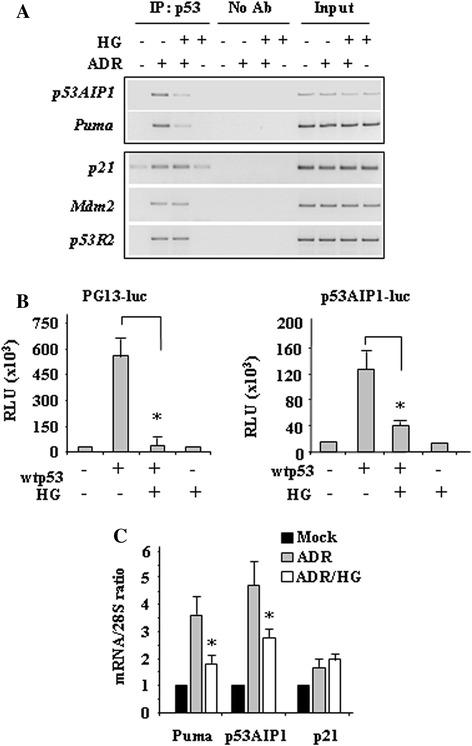
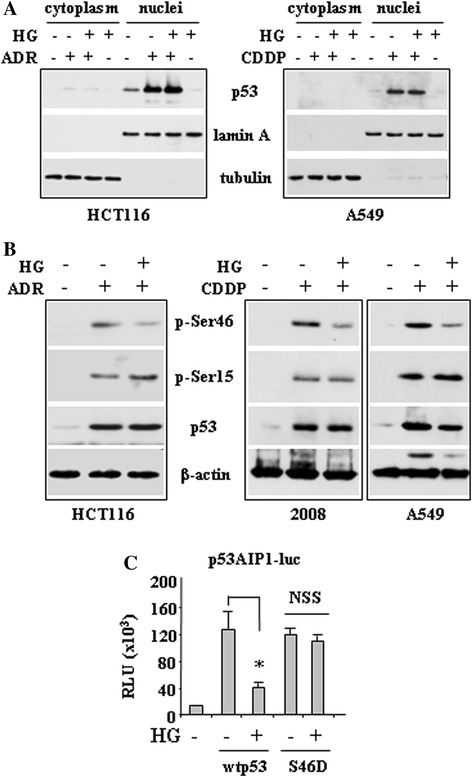
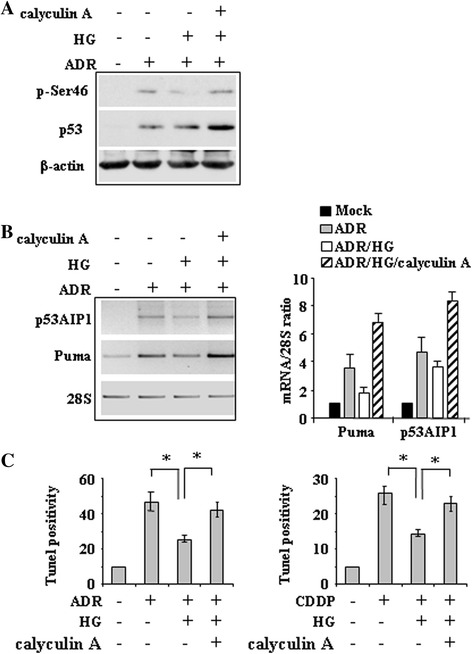
Methods and results: Here, we performed biochemical and molecular analyses in different cancer cell lines treated with chemotherapy in the presence or absence of high glucose condition. Analyses of p53 posttranslational modifications showed that drug-induced p53Ser46 phosphorylation was reduced by high glucose. Such reduction depended by high glucose-induced calyculin A-sensitive phosphatase(s), able to specifically target p53Ser46 phosphorylation. The specific effect on Ser46 phosphorylation was addressed by analysing Ser15 phosphorylation that instead was not modified by high glucose. In agreement, a constitutively phosphorylated Ser46D p53 mutant was resistant to high glucose. As a consequence of phosphoSer46 impairment, high glucose reduced the tumor cell response to drugs, correlating with reduced p53 apoptotic transactivation. The drug-induced apoptotic cell death, reduced by high glucose, was finally restored by the phosphatase inhibitor calyculin A.
Conclusions: These data indicate that high glucose specifically inhibited Ser46 phosphorylation thus reducing p53 apoptotic activity. These results uncover a new mechanism of p53 inactivation providing an interesting novel molecular link between metabolic diseases such as diabetes or obesity and tumor progression and resistance to therapies.
Figures
Similar articles
-
Overexpression of HIPK2 circumvents the blockade of apoptosis in chemoresistant ovarian cancer cells.Gynecol Oncol. 2008 Jun;109(3):403-10. doi: 10.1016/j.ygyno.2008.02.018. Epub 2008 Apr 18. Gynecol Oncol. 2008. PMID: 18395248
-
Inability of p53-reactivating compounds Nutlin-3 and RITA to overcome p53 resistance in tumor cells deficient in p53Ser46 phosphorylation.Biochem Biophys Res Commun. 2012 Jan 20;417(3):931-7. doi: 10.1016/j.bbrc.2011.11.161. Epub 2011 Dec 6. Biochem Biophys Res Commun. 2012. PMID: 22166212
-
HIPK2 modulates p53 activity towards pro-apoptotic transcription.Mol Cancer. 2009 Oct 14;8:85. doi: 10.1186/1476-4598-8-85. Mol Cancer. 2009. PMID: 19828042 Free PMC article.
-
Inhibitory role of cAMP on doxorubicin-induced apoptosis in pre-B ALL cells through dephosphorylation of p53 serine residues.Apoptosis. 2010 Feb;15(2):196-203. doi: 10.1007/s10495-009-0417-8. Apoptosis. 2010. PMID: 19882354
-
HIPK2-a therapeutical target to be (re)activated for tumor suppression: role in p53 activation and HIF-1α inhibition.Cell Cycle. 2010 Apr 1;9(7):1270-5. doi: 10.4161/cc.9.7.11125. Cell Cycle. 2010. PMID: 20234185 Review.
Cited by
-
Hyperglycemia during Adjuvant Chemotherapy as a Prognostic Factor in Breast Cancer Patients without Diabetes.J Breast Cancer. 2020 Aug;23(4):398-409. doi: 10.4048/jbc.2020.23.e44. J Breast Cancer. 2020. PMID: 32908790 Free PMC article.
-
ZnCl2 sustains the adriamycin-induced cell death inhibited by high glucose.Cell Death Dis. 2016 Jun 30;7(6):e2280. doi: 10.1038/cddis.2016.178. Cell Death Dis. 2016. PMID: 27362798 Free PMC article.
-
Hyperglycemia triggers HIPK2 protein degradation.Oncotarget. 2017 Jan 3;8(1):1190-1203. doi: 10.18632/oncotarget.13595. Oncotarget. 2017. PMID: 27901482 Free PMC article.
-
GPSD: a hybrid learning framework for the prediction of phosphatase-specific dephosphorylation sites.Brief Bioinform. 2024 Nov 22;26(1):bbae694. doi: 10.1093/bib/bbae694. Brief Bioinform. 2024. PMID: 39749667 Free PMC article.
-
The Role of Inflammation in Cancer: Mechanisms of Tumor Initiation, Progression, and Metastasis.Cells. 2025 Mar 25;14(7):488. doi: 10.3390/cells14070488. Cells. 2025. PMID: 40214442 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
