

Trans-Golgi network localized small GTPase RabA1d is involved in cell plate formation and oscillatory root hair growth
- PMID: 25260869
- PMCID: PMC4180857
- DOI: 10.1186/s12870-014-0252-0
Trans-Golgi network localized small GTPase RabA1d is involved in cell plate formation and oscillatory root hair growth
Abstract
Background: Small Rab GTPases are important regulators of vesicular trafficking in plants. AtRabA1d, a member of the RabA1 subfamily of small GTPases, was previously found in the vesicle-rich apical dome of growing root hairs suggesting a role during tip growth; however, its specific intracellular localization and role in plants has not been well described.
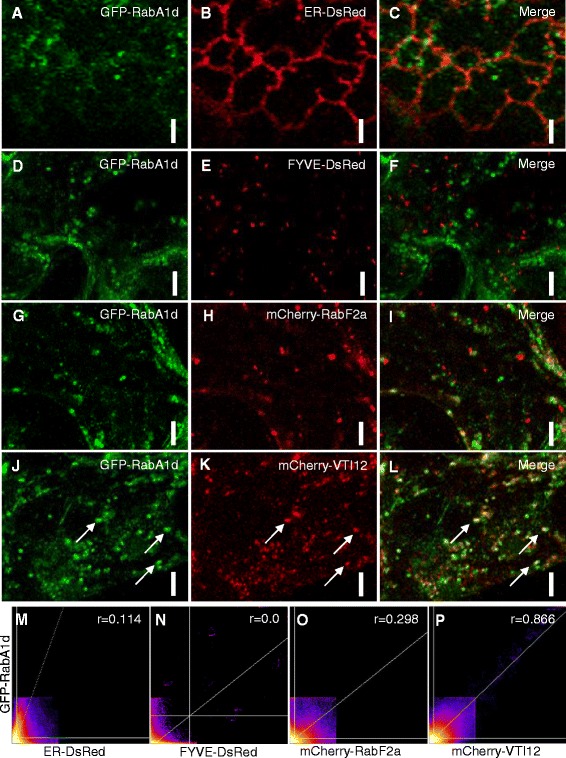
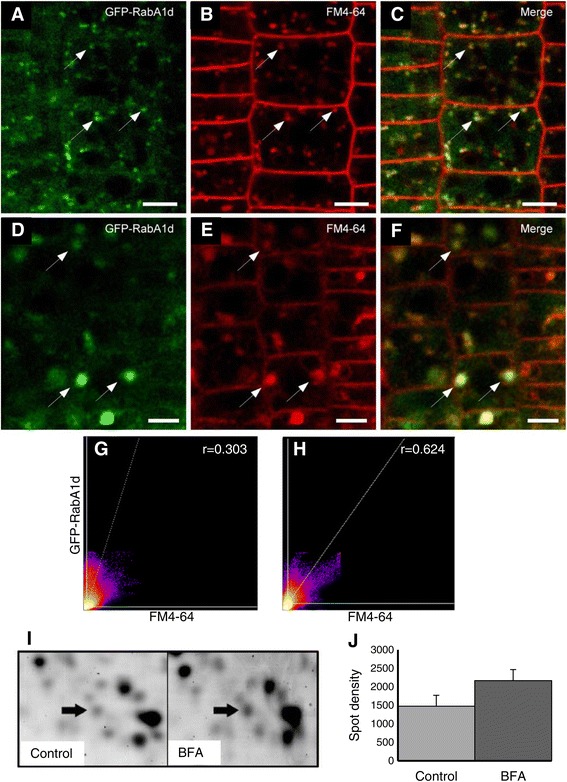
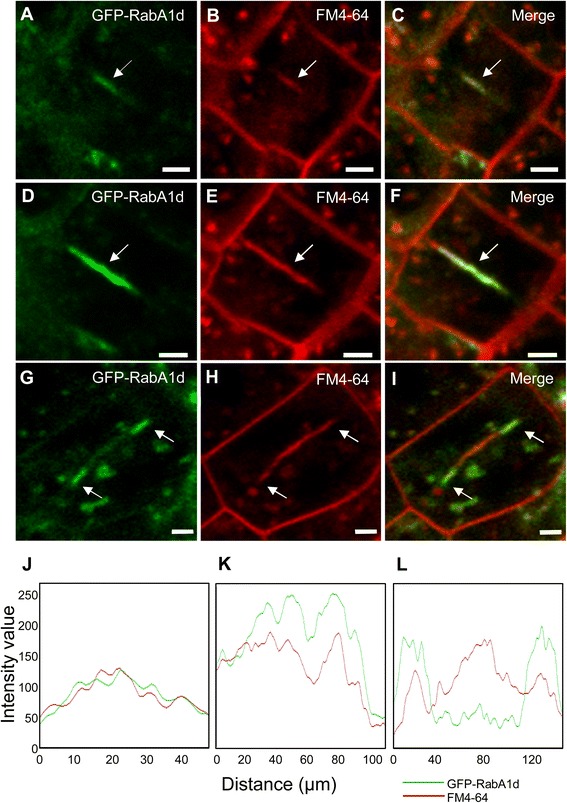
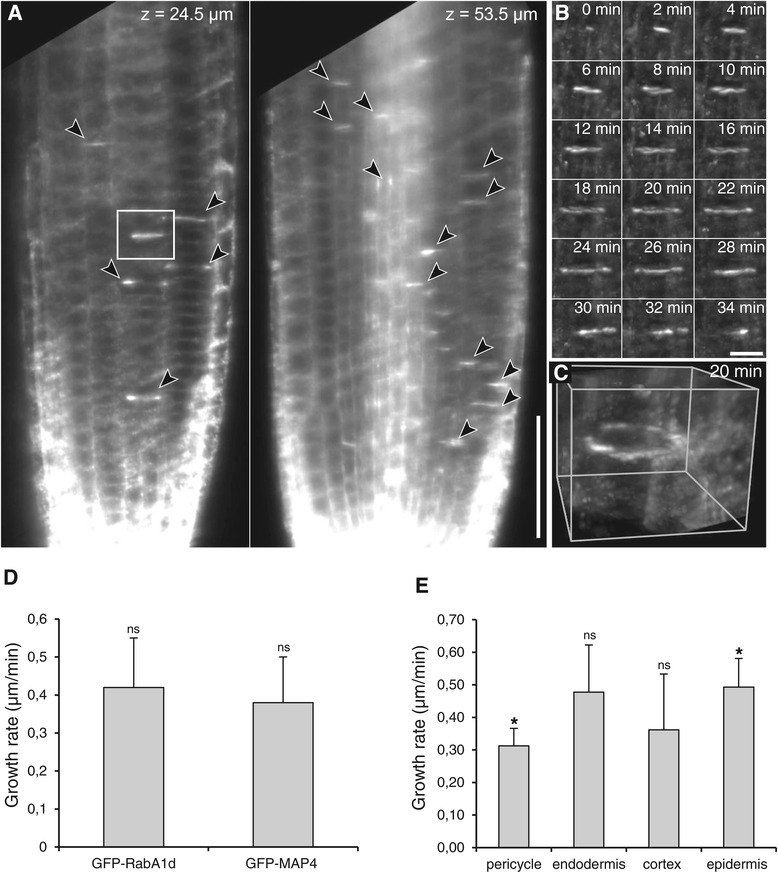
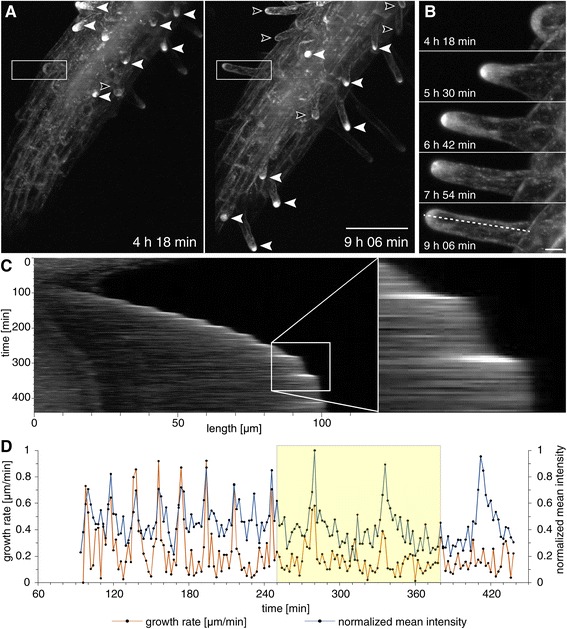
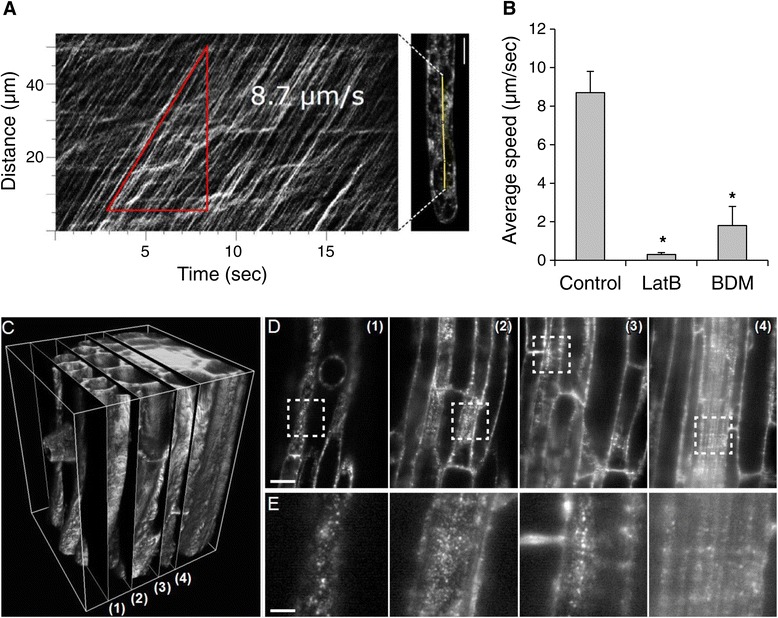
Results: The transient expression of 35S::GFP:RabA1d construct in Allium porrum and Nicotiana benthamiana revealed vesicular structures, which were further corroborated in stable transformed Arabidopsis thaliana plants. GFP-RabA1d colocalized with the trans-Golgi network marker mCherry-VTI12 and with early FM4-64-labeled endosomal compartments. Late endosomes and endoplasmic reticulum labeled with FYVE-DsRed and ER-DsRed, respectively, were devoid of GFP-RabA1d. The accumulation of GFP-RabA1d in the core of brefeldin A (BFA)-induced-compartments and the quantitative upregulation of RabA1d protein levels after BFA treatment confirmed the association of RabA1d with early endosomes/TGN and its role in vesicle trafficking. Light-sheet microscopy revealed involvement of RabA1d in root development. In root cells, GFP-RabA1d followed cell plate expansion consistently with cytokinesis-related vesicular trafficking and membrane recycling. GFP-RabA1d accumulated in disc-like structures of nascent cell plates, which progressively evolved to marginal ring-like structures of the growing cell plates. During root hair growth and development, GFP-RabA1d was enriched at root hair bulges and at the apical dome of vigorously elongating root hairs. Importantly, GFP-RabA1d signal intensity exhibited an oscillatory behavior in-phase with tip growth. Progressively, this tip localization dissapeared in mature root hairs suggesting a link between tip localization of RabA1d and root hair elongation. Our results support a RabA1d role in events that require vigorous membrane trafficking.
Conclusions: RabA1d is located in early endosomes/TGN and is involved in vesicle trafficking. RabA1d participates in both cell plate formation and root hair oscillatory tip growth. The specific GFP-RabA1d subcellular localization confirms a correlation between its specific spatio-temporal accumulation and local vesicle trafficking requirements during cell plate and root hair formation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
