

The high molecular weight neurofilament subunit plays an essential role in axonal outgrowth and stabilization
- PMID: 25260918
- PMCID: PMC4197446
- DOI: 10.1242/bio.20149779
The high molecular weight neurofilament subunit plays an essential role in axonal outgrowth and stabilization
Abstract
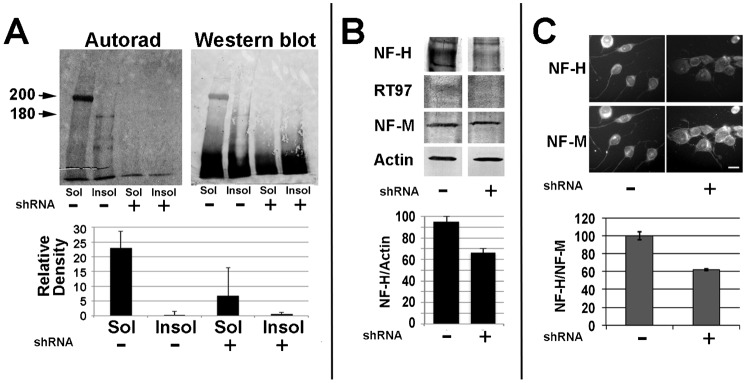
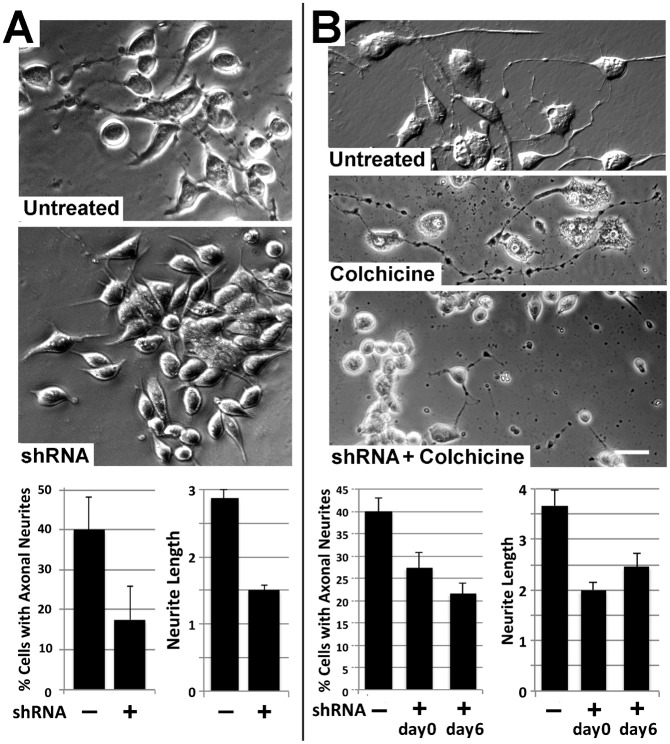
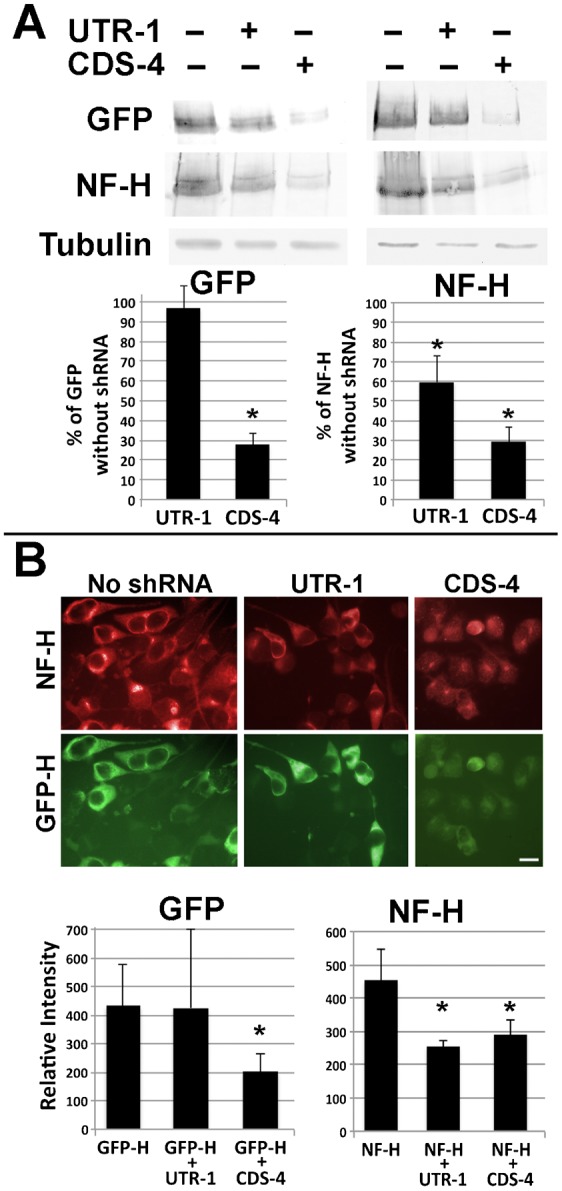
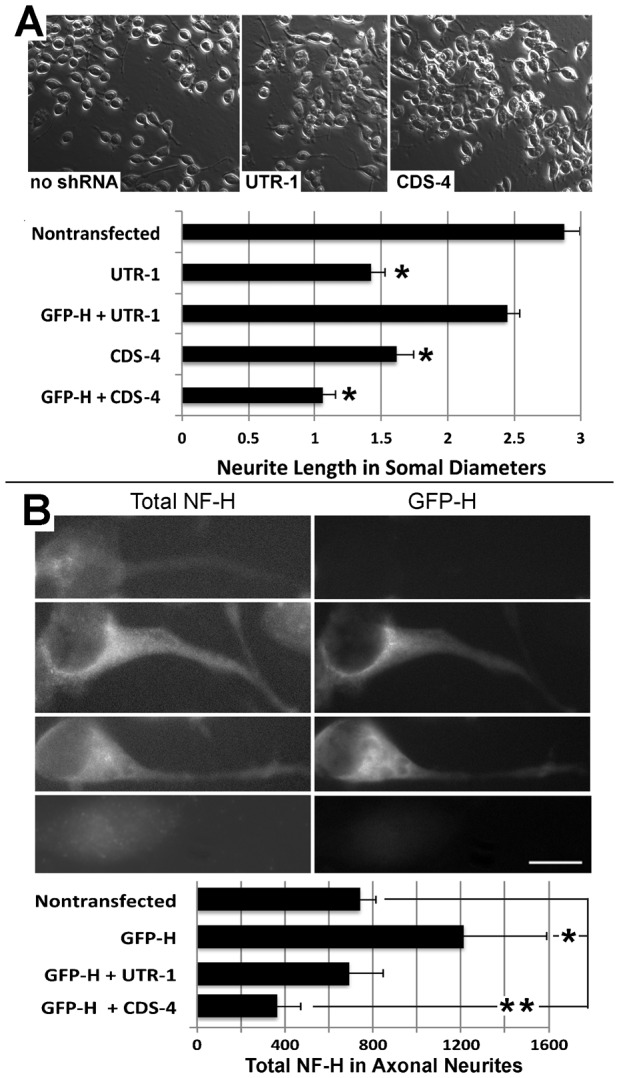
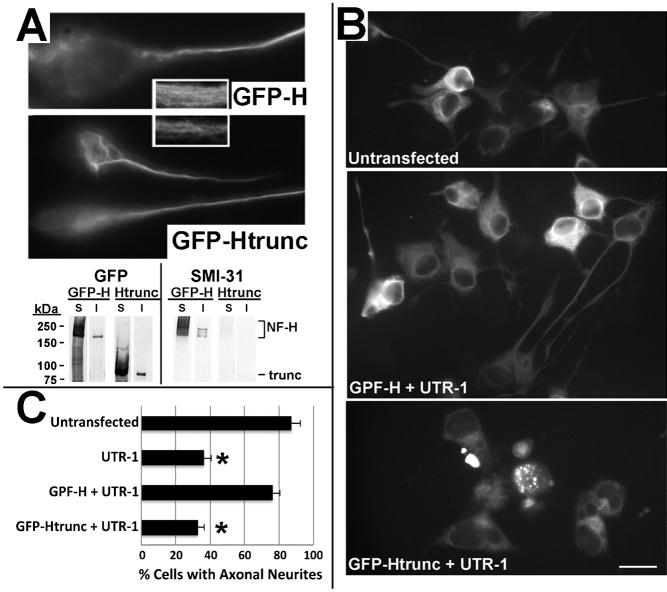
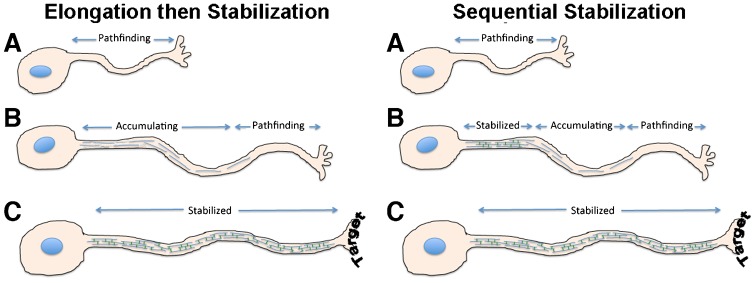
Neurofilaments (NFs) are thought to provide structural support to mature axons via crosslinking of cytoskeletal elements mediated by the C-terminal region of the high molecular weight NF subunit (NF-H). Herein, we inhibited NF-H expression in differentiating mouse NB2a/d1 cells with shRNA directed against murine NF-H without affecting other NF subunits, microtubules or actin. shRNA-mediated NF-H knockdown not only in compromised of late-stage axonal neurite stabilization but also compromised early stages of axonal neurite elongation. Expression of exogenous rat NF-H was able to compensate for knockdown of endogenous NF-H and restored the development and stabilization of axonal neurites. This rescue was prevented by simultaneous treatment with shRNA that inhibited both rat and murine NF-H, or by expression of exogenous rat NF-H lacking the C-terminal sidearm during knockdown of endogenous NF-H. Demonstration of a role for NF-H in the early stages of axonal elaboration suggests that axonal stabilization is not delayed until synaptogenesis, but rather that the developing axon undergoes sequential NF-H-mediated stabilization along its length in a proximal-distal manner, which supports continued pathfinding in distal, unstabilized regions.
Keywords: Axonal outgrowth; Axonal stability; Cytoskeleton; Nervous system development; Neurofilament.
© 2014. Published by The Company of Biologists Ltd.
Conflict of interest statement
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
