

Jasmonate-dependent induction of polyphenol oxidase activity in tomato foliage is important for defense against Spodoptera exigua but not against Manduca sexta
- PMID: 25261073
- PMCID: PMC4189532
- DOI: 10.1186/s12870-014-0257-8
Jasmonate-dependent induction of polyphenol oxidase activity in tomato foliage is important for defense against Spodoptera exigua but not against Manduca sexta
Abstract
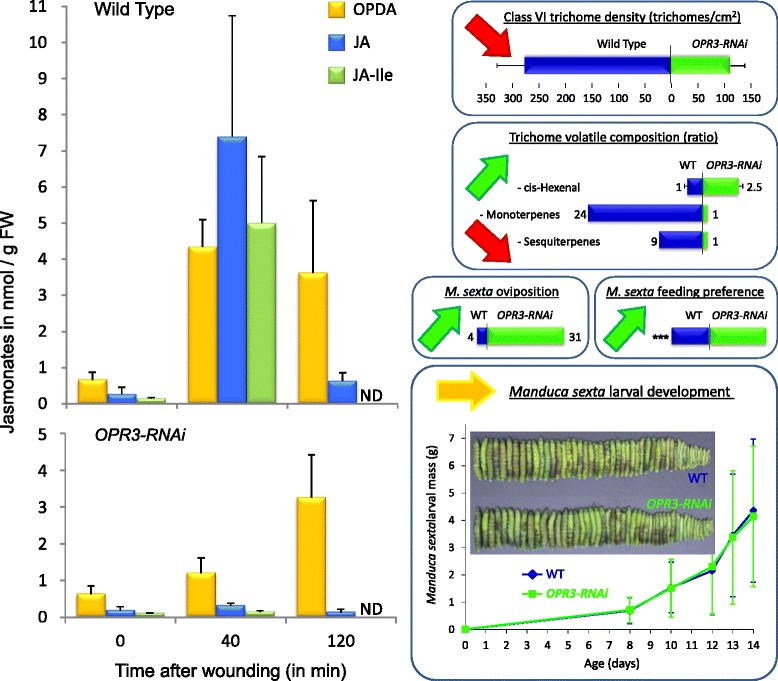
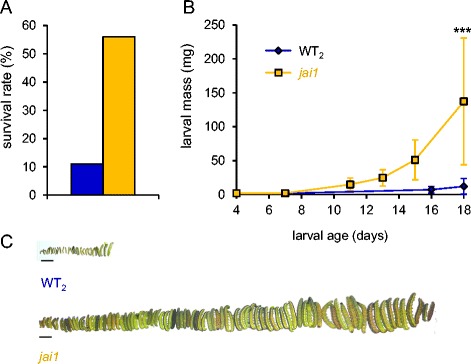
Background: Jasmonates are involved in plant defense, participating in the timely induction of defense responses against insect herbivores from different feeding guilds and with different degrees of host specialization. It is less clear to what extent the induction of plant defense is controlled by different members of the jasmonate family and how specificity of the response is achieved. Using transgenic plants blocked in jasmonic acid (JA) biosynthesis, we previously showed that JA is required for the formation of glandular trichomes and trichome-borne metabolites as constitutive defense traits in tomato, affecting oviposition and feeding behavior of the specialist Manduca sexta. In contrast, JA was not required for the local induction of defense gene expression after wounding. In JA-deficient plants, the JA precursor oxophytodienoic acid (OPDA) substituted as a regulator of defense gene expression maintaining considerable resistance against M. sexta larvae. In this study, we investigate the contribution of JA and OPDA to defense against the generalist herbivore Spodoptera exigua.
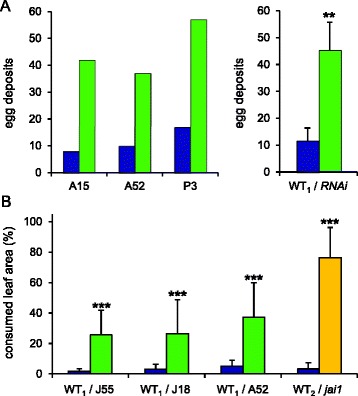
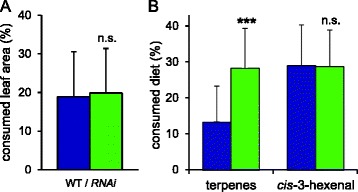
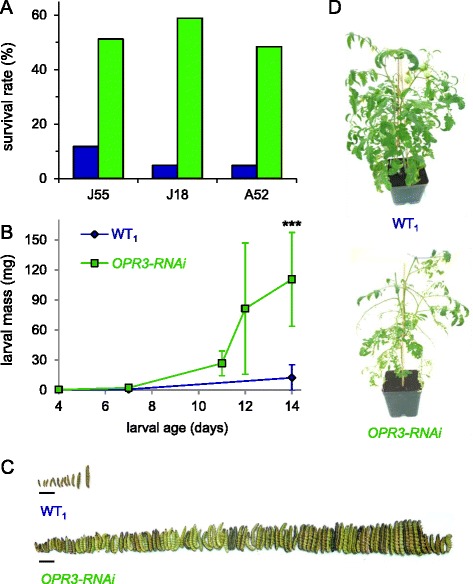
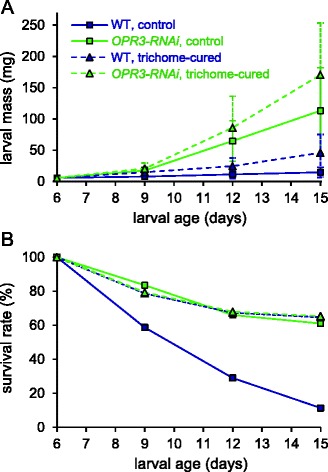
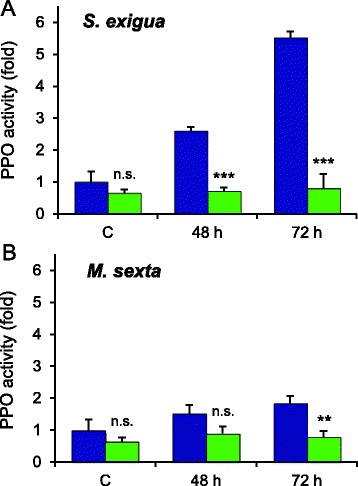
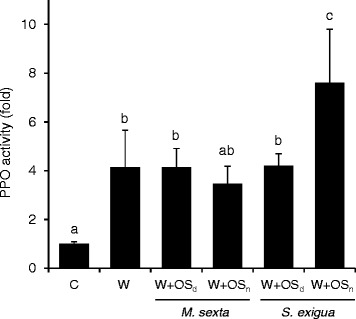
Results: S. exigua preferred JA-deficient over wild-type tomato plants as a host for both oviposition and feeding. Feeding preference for JA-deficient plants was caused by constitutively reduced levels of repellent terpenes. Growth and development of the larvae, on the other hand, were controlled by additional JA-dependent defense traits, including the JA-mediated induction of foliar polyphenol oxidase (PPO) activity. PPO induction was more pronounced after S. exigua herbivory as compared to mechanical wounding or M. sexta feeding. The difference was attributed to an elicitor exclusively present in S. exigua oral secretions.
Conclusions: The behavior of M. sexta and S. exigua during oviposition and feeding is controlled by constitutive JA/JA-Ile-dependent defense traits involving mono- and sesquiterpenes in both species, and cis-3-hexenal as an additional chemical cue for M. sexta. The requirement of jasmonates for resistance of tomato plants against caterpillar feeding differs for the two species. While the OPDA-mediated induction of local defense is sufficient to restrict growth and development of M. sexta larvae in absence of JA/JA-Ile, defense against S. exigua relied on additional JA/JA-Ile dependent factors, including the induction of foliar polyphenol oxidase activity in response to S. exigua oral secretions.
Figures
References
-
- Ehrlich PR, Raven PH. Butterflies and plants: a study in coevolution. Ecology. 1964;18:586–608.
-
- Janz N. Ehrlich and Raven revisited: Mechanisms underlying codiversification of plants and enemies. Ann Rev Ecol Evol Sys. 2011;42:71–89. doi: 10.1146/annurev-ecolsys-102710-145024. - DOI
-
- Walling LL. The myriad plant responses to herbivores. J Plant Growth Regul. 2000;19:195–216. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
