

Enzyme clustering accelerates processing of intermediates through metabolic channeling
- PMID: 25262299
- PMCID: PMC4666537
- DOI: 10.1038/nbt.3018
Enzyme clustering accelerates processing of intermediates through metabolic channeling
Abstract
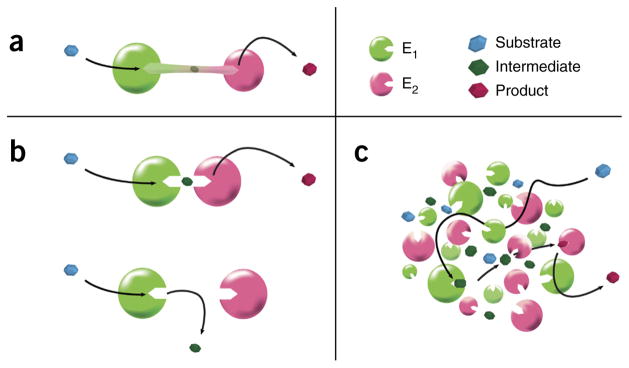
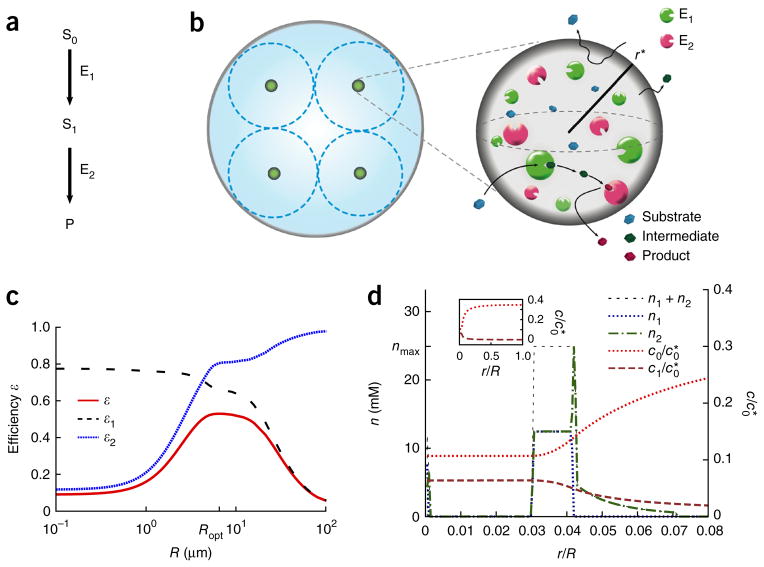
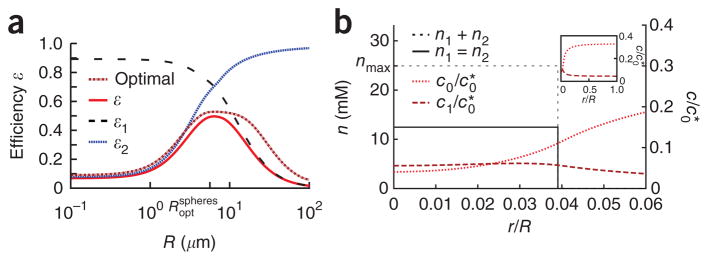
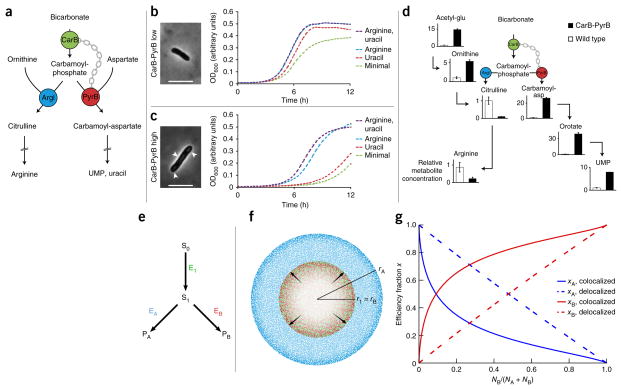
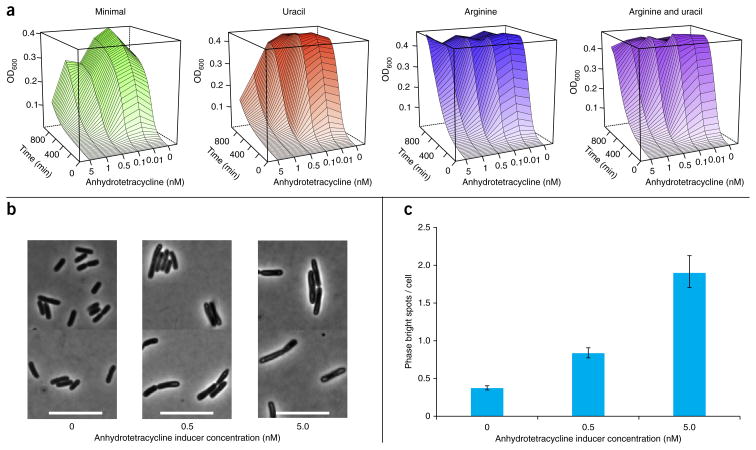
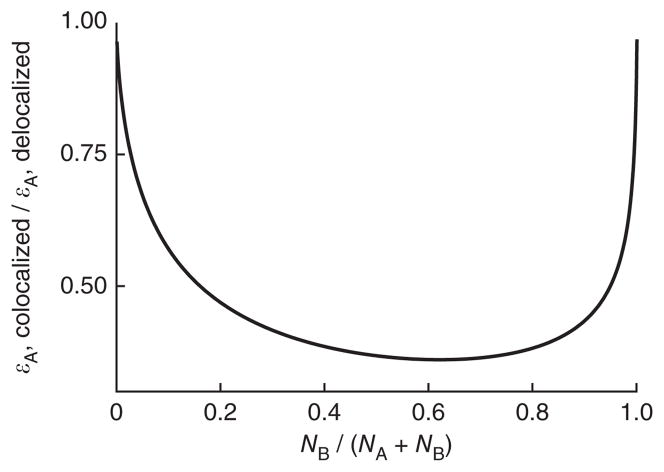
We present a quantitative model to demonstrate that coclustering multiple enzymes into compact agglomerates accelerates the processing of intermediates, yielding the same efficiency benefits as direct channeling, a well-known mechanism in which enzymes are funneled between enzyme active sites through a physical tunnel. The model predicts the separation and size of coclusters that maximize metabolic efficiency, and this prediction is in agreement with previously reported spacings between coclusters in mammalian cells. For direct validation, we study a metabolic branch point in Escherichia coli and experimentally confirm the model prediction that enzyme agglomerates can accelerate the processing of a shared intermediate by one branch, and thus regulate steady-state flux division. Our studies establish a quantitative framework to understand coclustering-mediated metabolic channeling and its application to both efficiency improvement and metabolic regulation.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- James CL, Viola RE. Production and characterization of bifunctional enzymes. Substrate channeling in the aspartate pathway. Biochemistry. 2002;41:3726–3731. - PubMed
-
- Dunn MF, et al. The tryptophan synthase bienzyme complex transfers indole between the alpha and beta sites via a 25–30 ANG long tunnel. Biochemistry. 1990;29:8598–8607. - PubMed
-
- Huang X, Holden HM, Raushel FM. Channeling of substrates and intermediates in enzyme-catalyzed reactions. Annu Rev Biochem. 2001;70:149–180. - PubMed
-
- Welch GR. On the role of organized multienzyme systems in cellular metabolism: A general synthesis. Prog Biophys Mol Biol. 1978;32:103–191. - PubMed
-
- Bernhard SA, Srivastava DK. Direct transfer of metabolites via enzyme-enzyme complexes: evidence and physiological significance. NATO ASI Series. 1987;127:143–163.
