

Quantitative and in toto imaging in ascidians: working toward an image-centric systems biology of chordate morphogenesis
- PMID: 25262824
- PMCID: PMC4378666
- DOI: 10.1002/dvg.22828
Quantitative and in toto imaging in ascidians: working toward an image-centric systems biology of chordate morphogenesis
Abstract
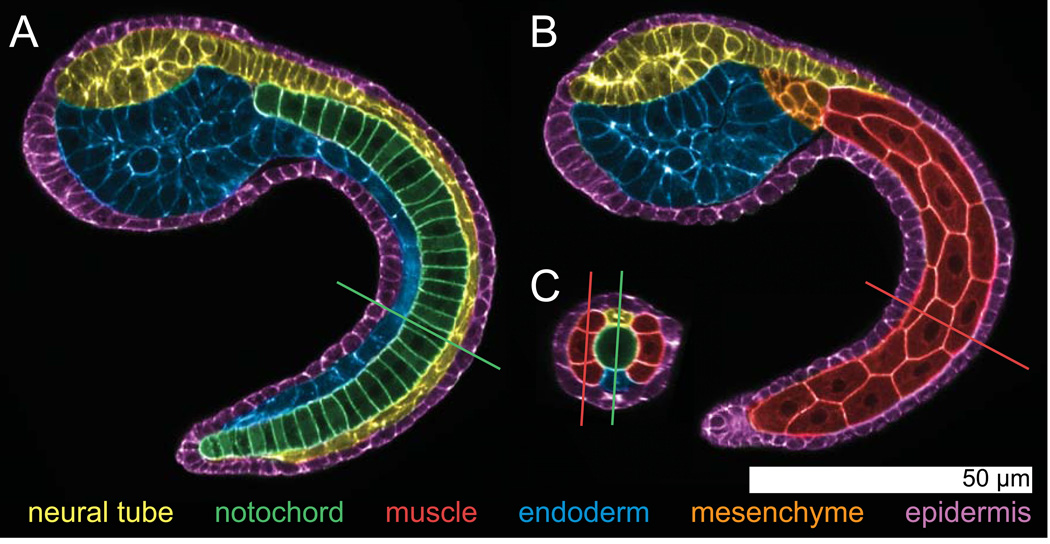
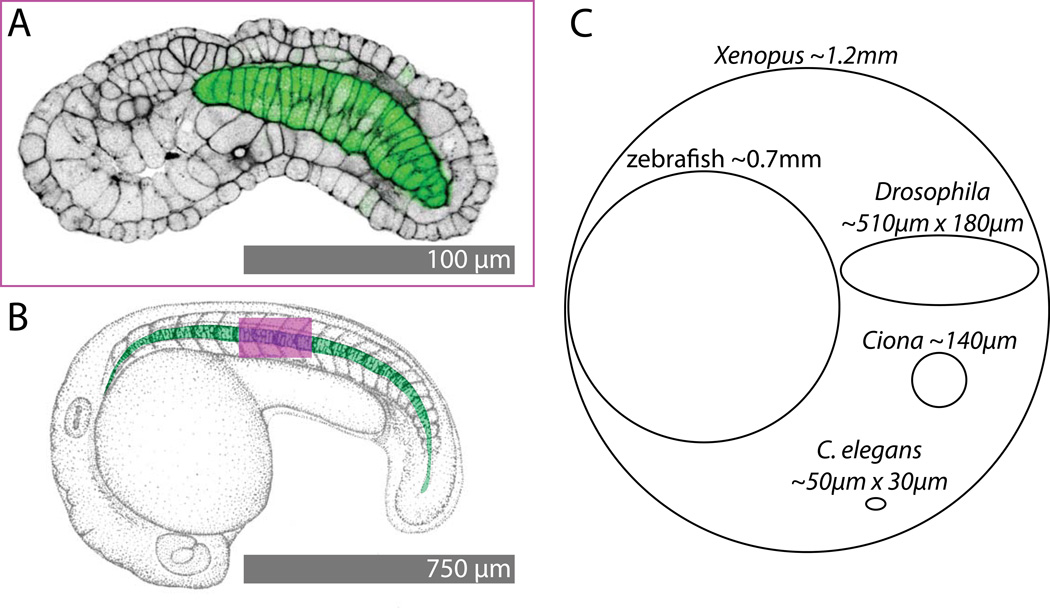
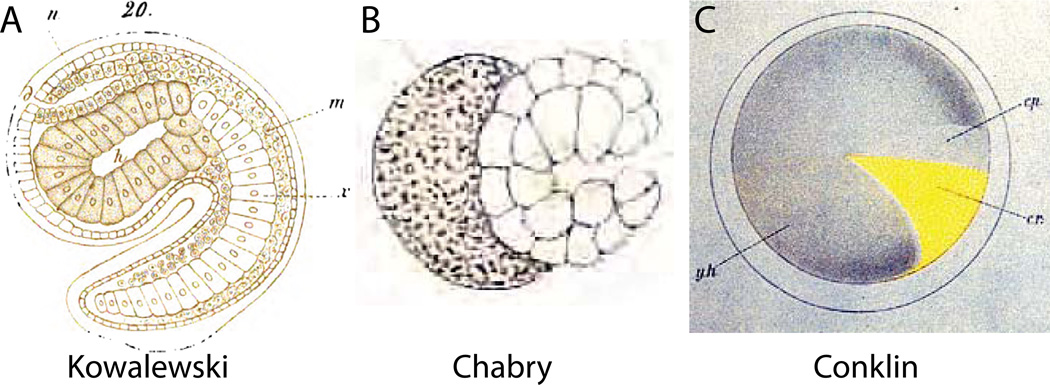
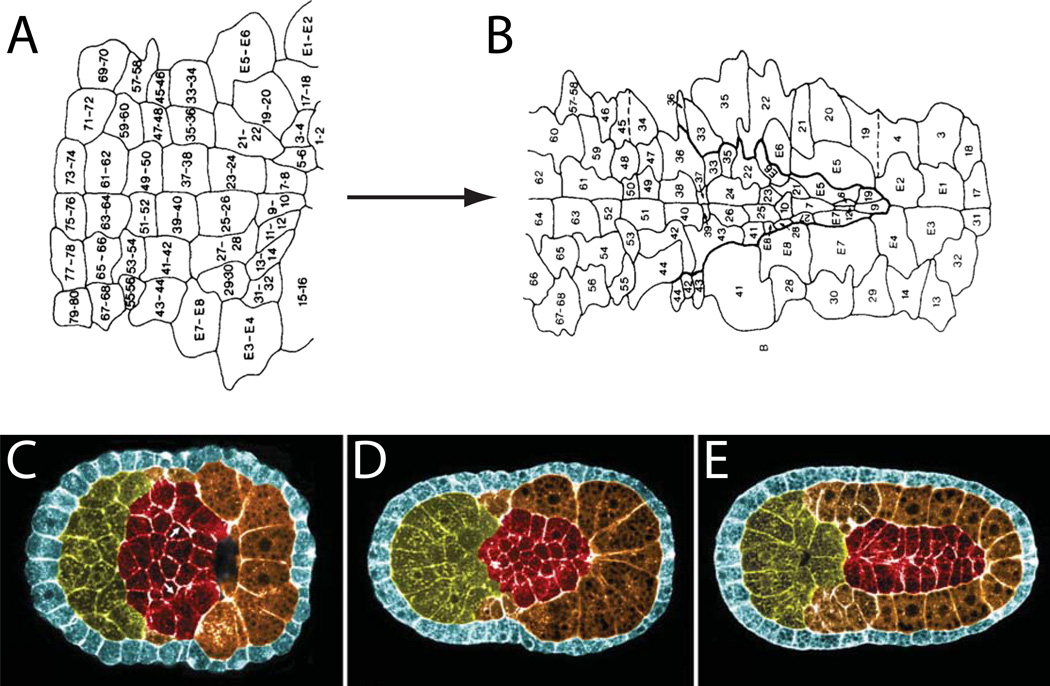
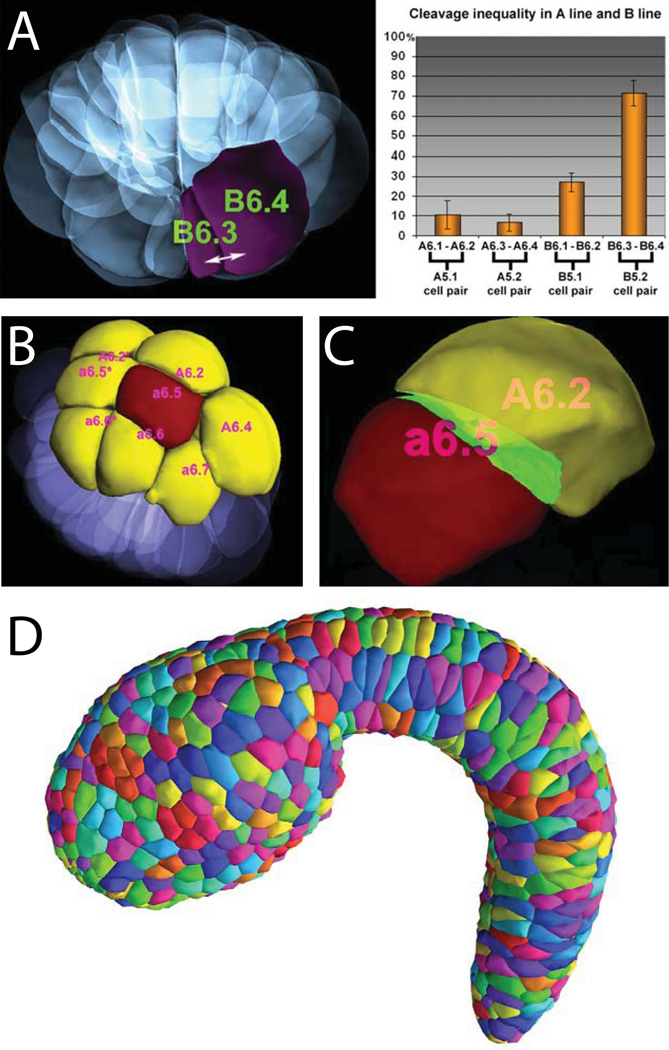
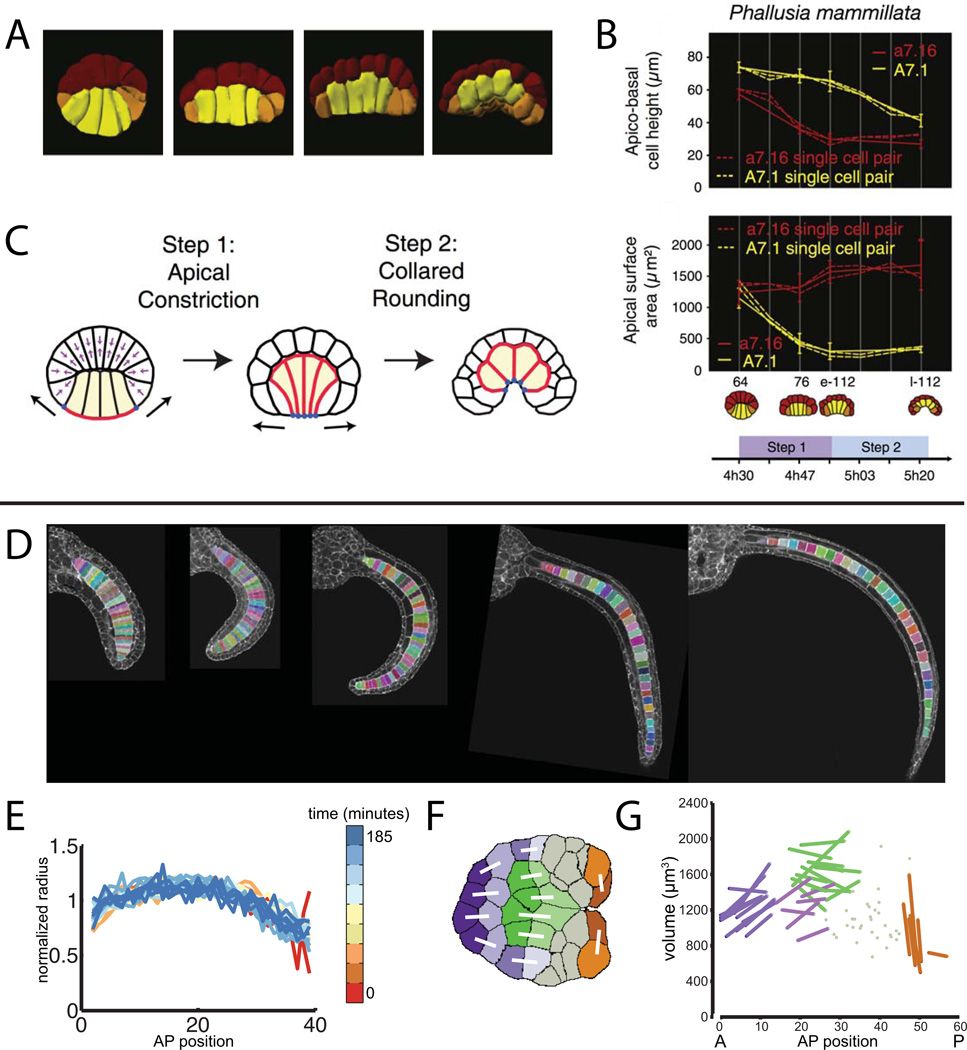
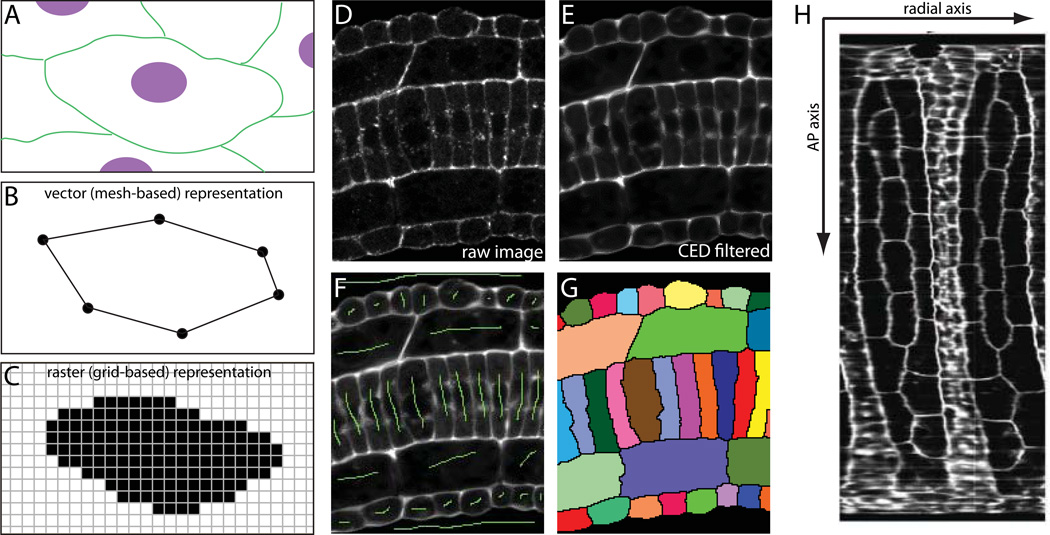
Developmental biology relies heavily on microscopy to image the finely controlled cell behaviors that drive embryonic development. Most embryos are large enough that a field of view with the resolution and magnification needed to resolve single cells will not span more than a small region of the embryo. Ascidian embryos, however, are sufficiently small that they can be imaged in toto with fine subcellular detail using conventional microscopes and objectives. Unlike other model organisms with particularly small embryos, ascidians have a chordate embryonic body plan that includes a notochord, hollow dorsal neural tube, heart primordium and numerous other anatomical details conserved with the vertebrates. Here we compare the size and anatomy of ascidian embryos with those of more traditional model organisms, and relate these features to the capabilities of both conventional and exotic imaging methods. We review the emergence of Ciona and related ascidian species as model organisms for a new era of image-based developmental systems biology. We conclude by discussing some important challenges in ascidian imaging and image analysis that remain to be solved.
Keywords: Ciona; ascidian; cell segmentation; in toto imaging; morphogenesis.
© 2014 Wiley Periodicals, Inc.
Figures
References
-
- Amos WB, White JG. How the confocal laser scanning microscope entered biological research. Biol Cell. 2003;95:335–342. - PubMed
-
- Begasse ML, Hyman AA. The first cell cycle of the Caenorhabditis elegans embryo: spatial and temporal control of an asymmetric cell division. Results Probl Cell Differ. 2011;53:109–133. - PubMed
-
- Brown DD. A tribute to the Xenopus laevis oocyte and egg. J Biol Chem. 2004;279:45291–45299. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
