

Polybivalency and disordered proteins in ordering macromolecular assemblies
- PMID: 25263009
- PMCID: PMC4339520
- DOI: 10.1016/j.semcdb.2014.09.016
Polybivalency and disordered proteins in ordering macromolecular assemblies
Abstract
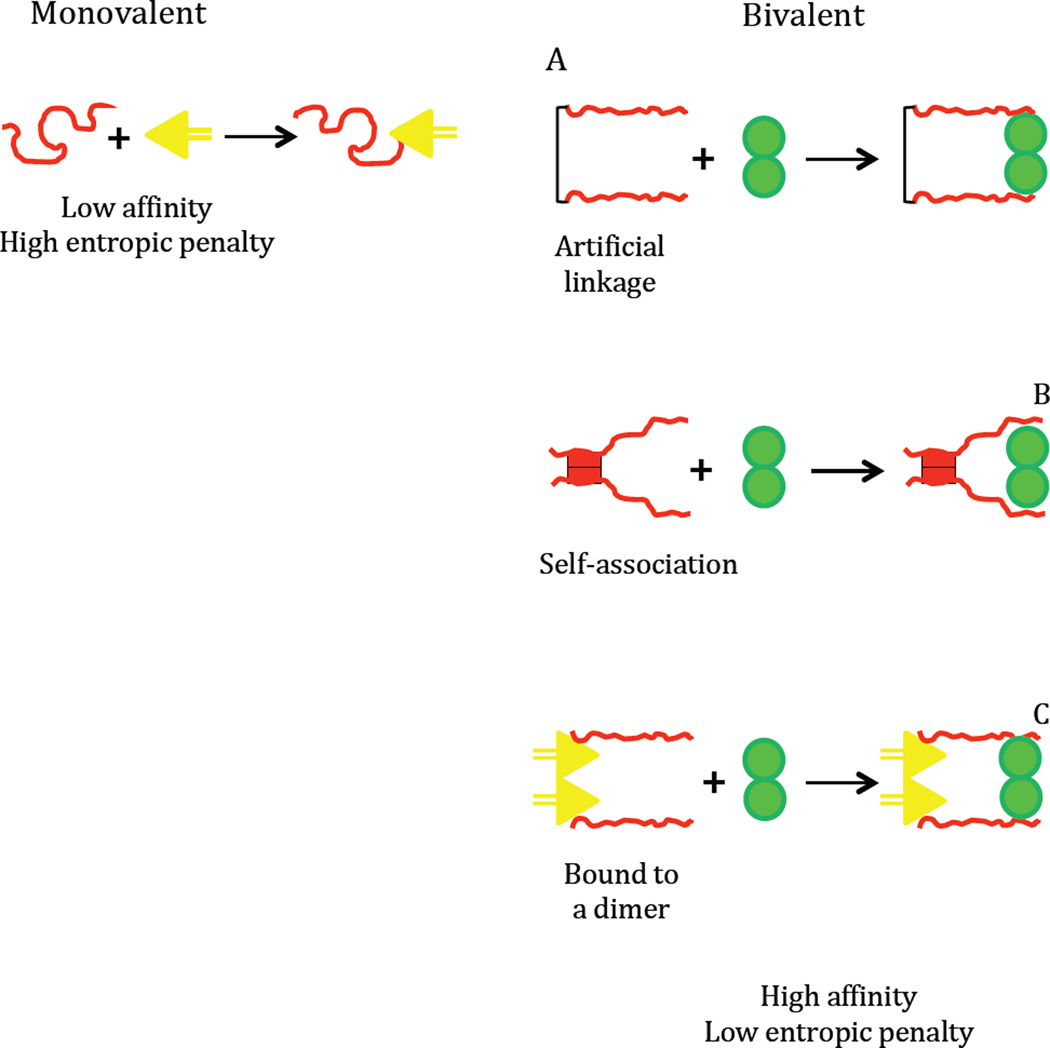
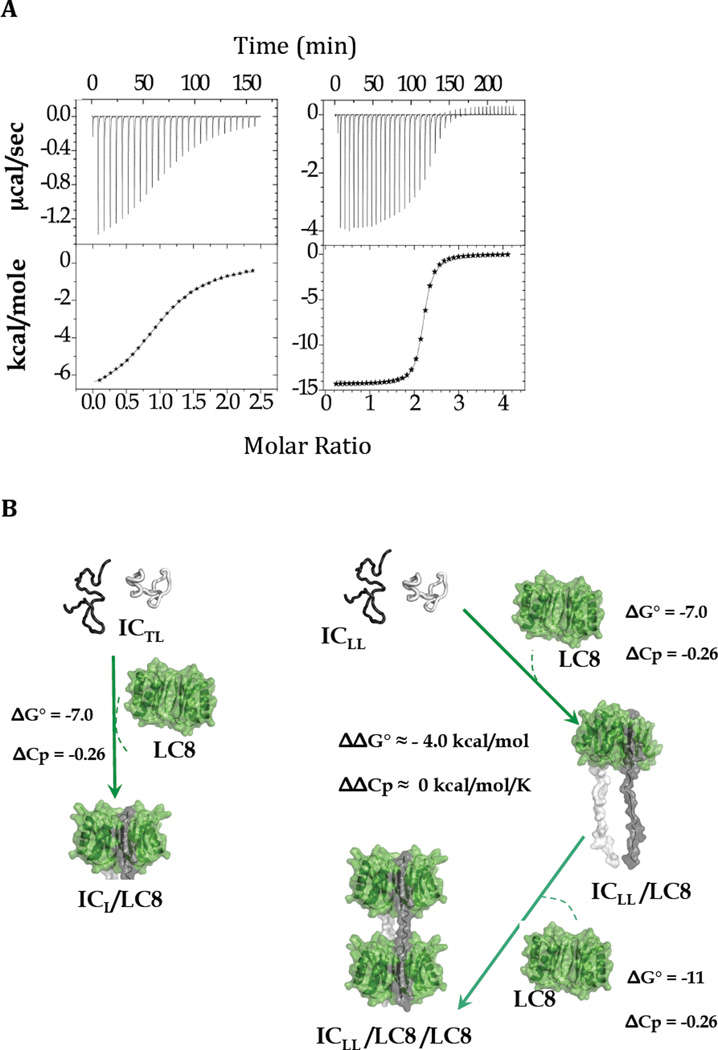
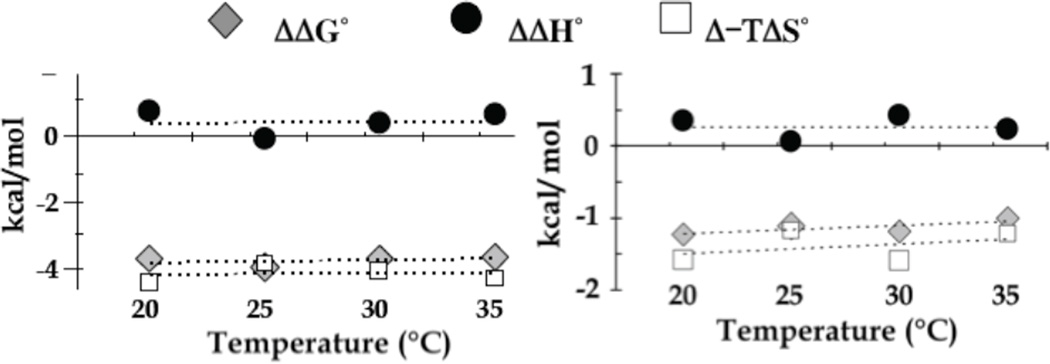
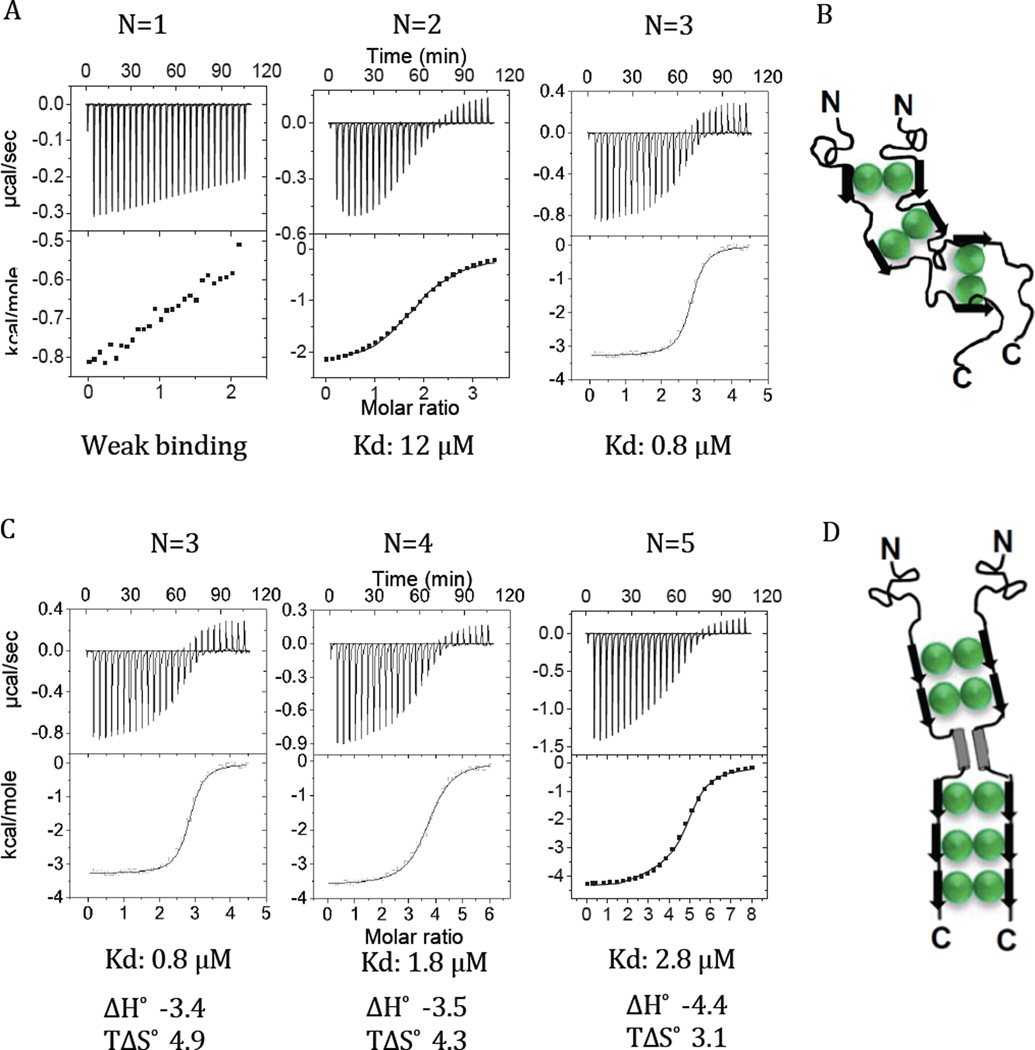
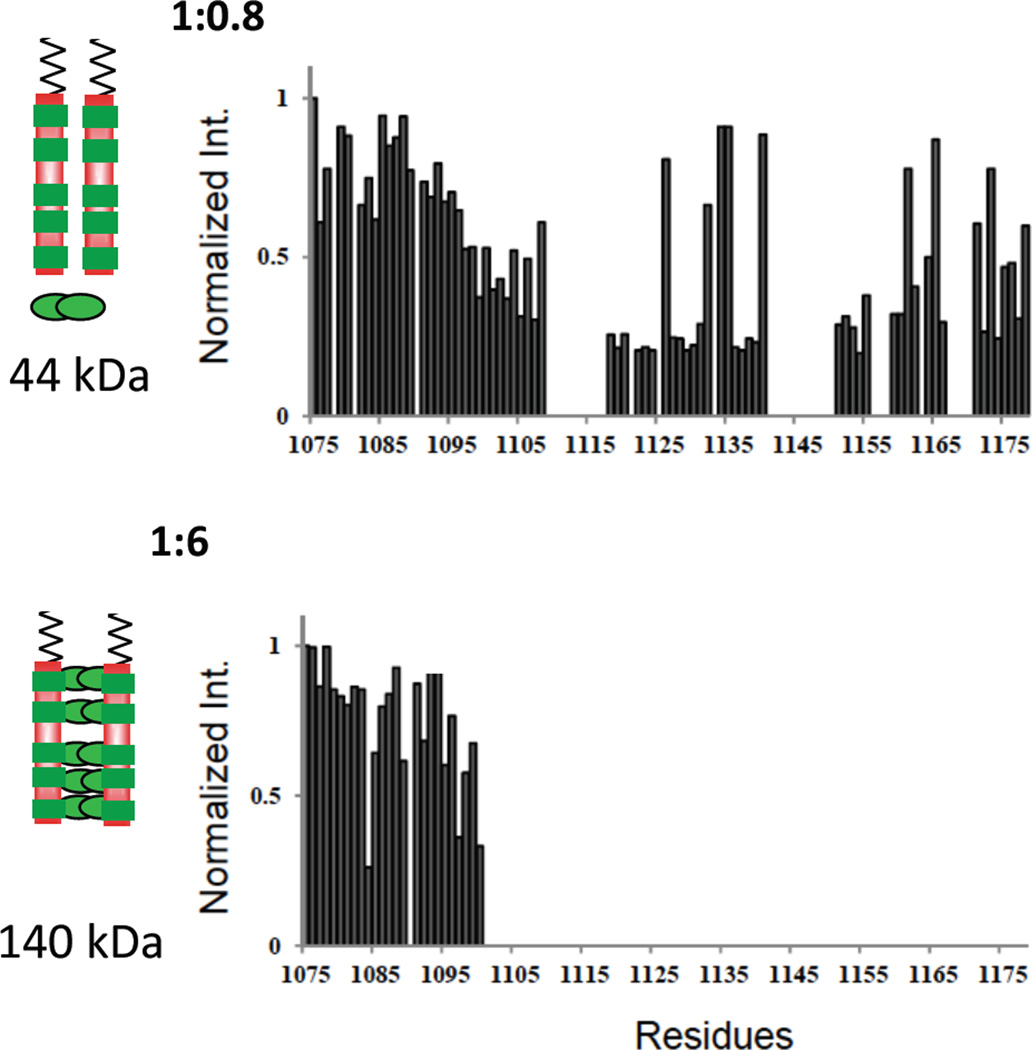
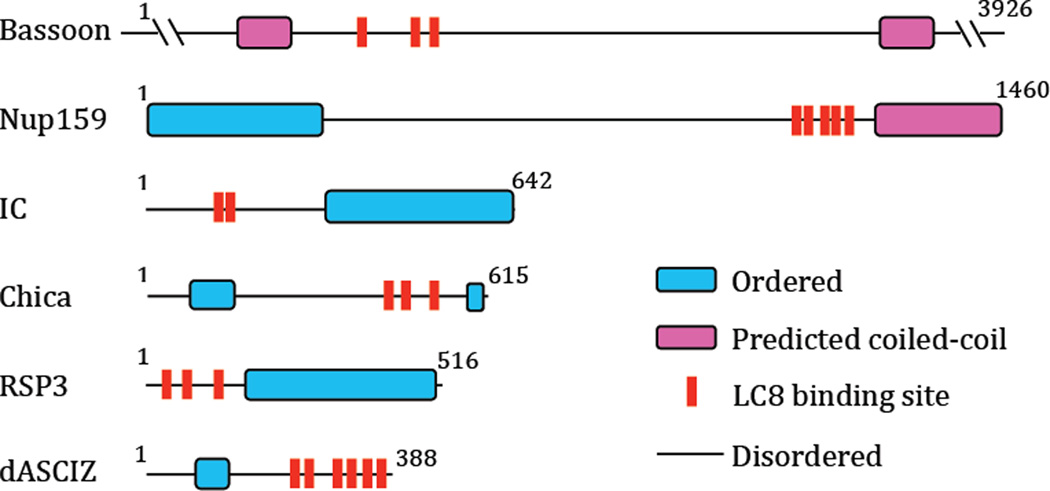
Intrinsically disordered proteins (IDPs) are prevalent in macromolecular assemblies and are thought to mediate protein recognition in complex regulatory processes and signaling pathways. The formation of a polybivalent scaffold is a key process by which IDPs drive early steps in macromolecular assemblies. Three intrinsically disordered proteins, IC, Swallow and Nup159, are core components, respectively, of cytoplasmic dynein, bicoid mRNA localization apparatus, and nuclear pore complexes. In all three systems, the hub protein LC8 recognizes on the IDP, short linear motifs that are fully disordered in the apo form, but adopt a β-strand when bound to LC8. The IDP/LC8 complex forms a bivalent scaffold primed to bind additional bivalent ligands. Scaffold formation also promotes self-association and/or higher order organization of the IDP components at a site distant from LC8 binding. Rigorous thermodynamic analyses imply that association of additional bivalent ligands is driven by entropic effects where the first binding event is weak but subsequent binding of additional ligands occurs with higher affinity. Here, we review specific examples of macromolecular assemblies in which polybivalency of aligned IDP duplexes not only enhances binding affinity and results in formation of a stable complex but also compensates unfavorable steric and enthalpic interactions. We propose that polybivalent scaffold assembly involving IDPs and LC8-like proteins is a general process in the cell biology of a class of multi-protein structures that are stable yet fine-tuned for diverse cellular requirements.
Keywords: Bivalency; Enthalpy–entropy compensation; Intrinsically disordered proteins; LC8; Macromolecular assembly; Poly-bivalent scaffold.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Benison G, Nyarko A, Barbar E. Heteronuclear NMR identifies a nascent helix in intrinsically disordered dynein intermediate chain: implications for folding and dimerization. J Mol Biol. 2006;362:1082–1093. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
