

Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine
- PMID: 25263219
- PMCID: PMC4260654
- DOI: 10.1016/j.chom.2014.09.001
Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine
Abstract
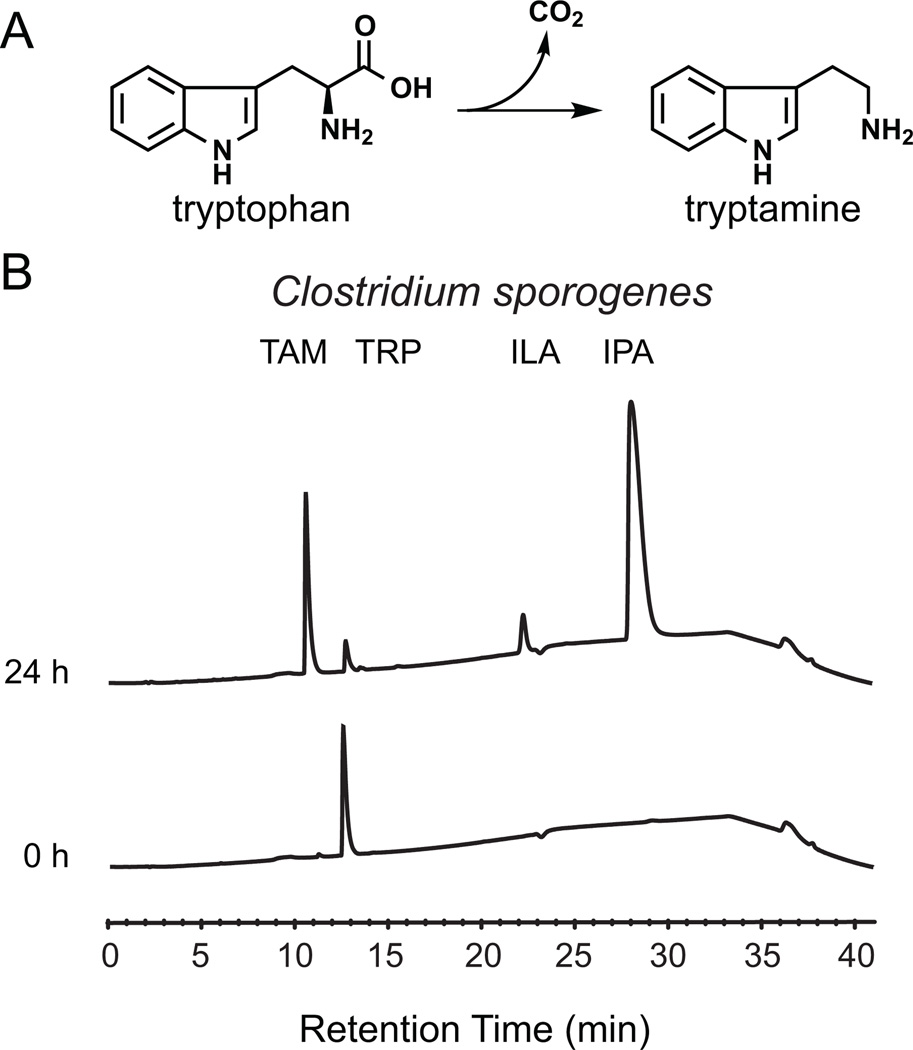
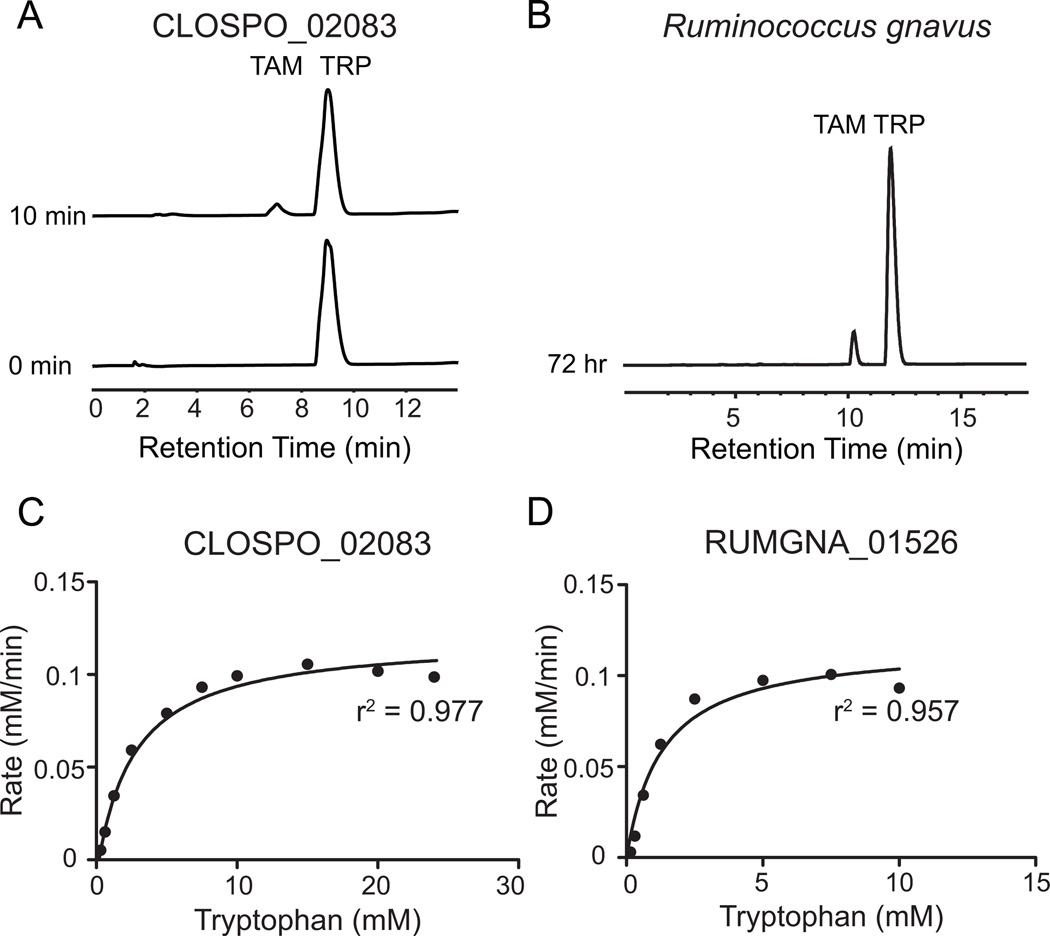
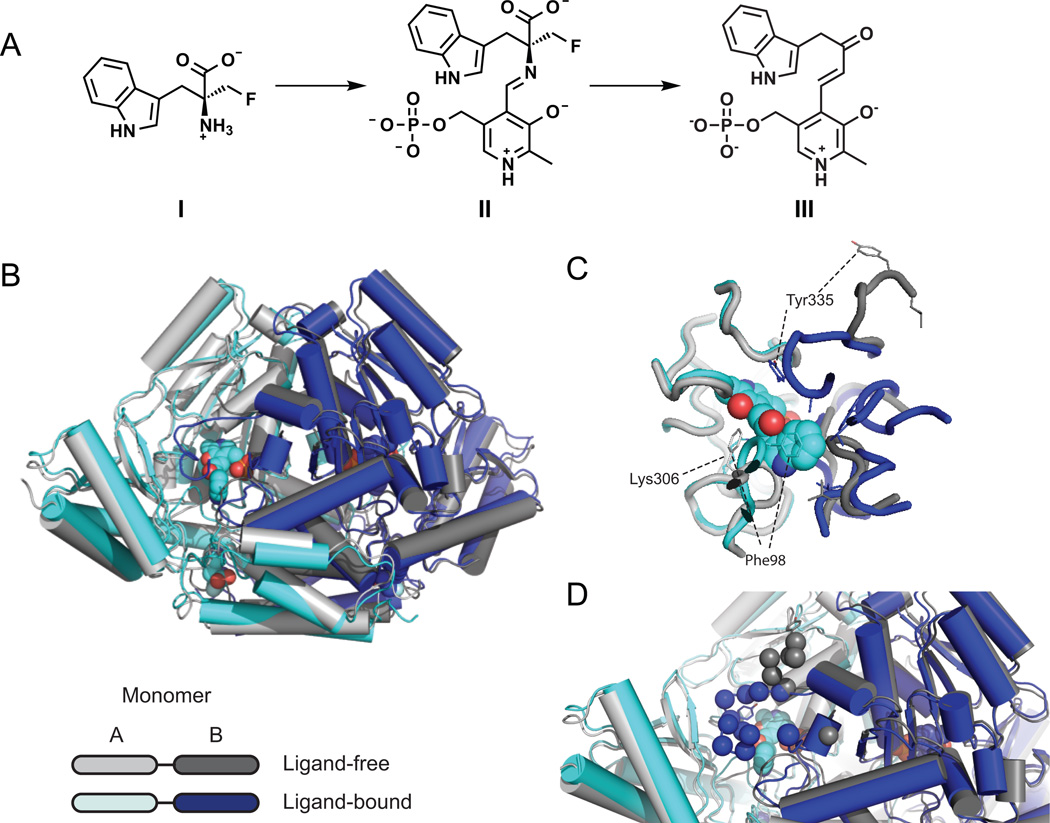
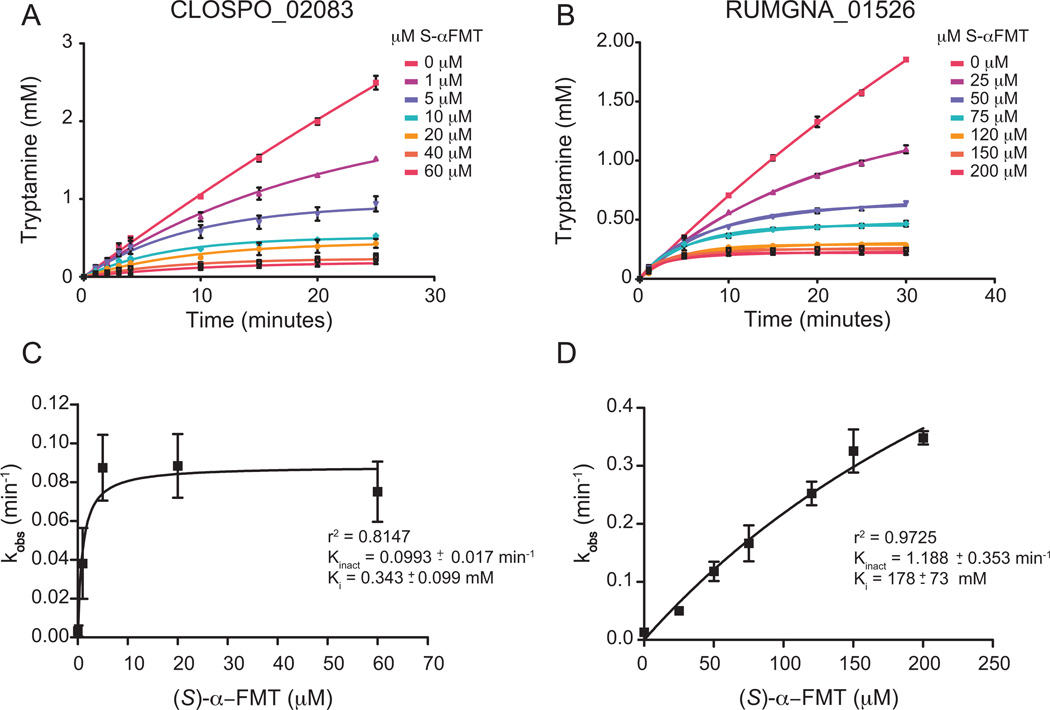
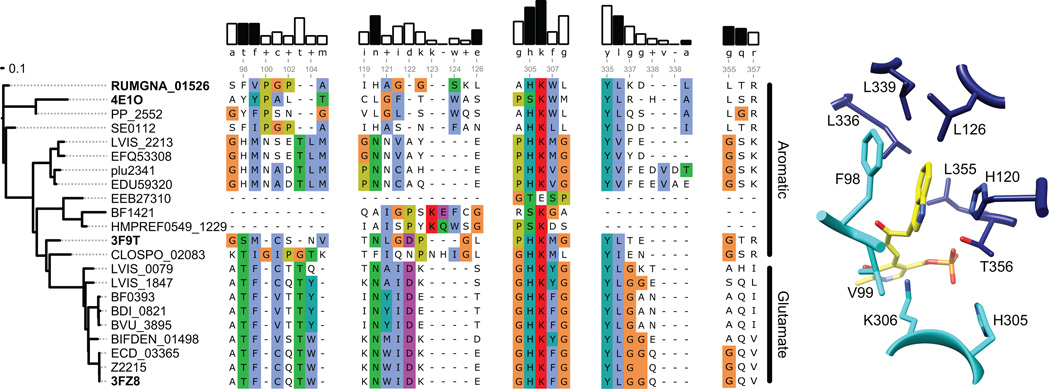
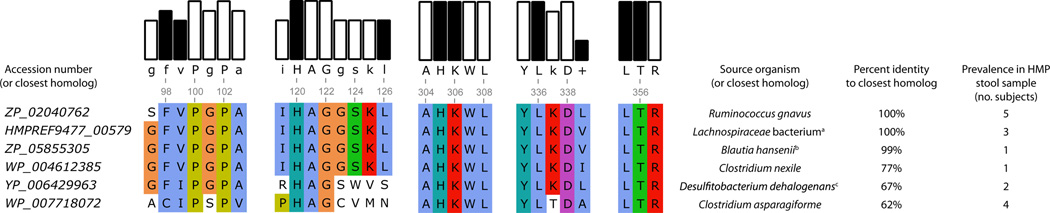
Several recent studies describe the influence of the gut microbiota on host brain and behavior. However, the mechanisms responsible for microbiota-nervous system interactions are largely unknown. Using a combination of genetics, biochemistry, and crystallography, we identify and characterize two phylogenetically distinct enzymes found in the human microbiome that decarboxylate tryptophan to form the β-arylamine neurotransmitter tryptamine. Although this enzymatic activity is exceedingly rare among bacteria more broadly, analysis of the Human Microbiome Project data demonstrate that at least 10% of the human population harbors at least one bacterium encoding a tryptophan decarboxylase in their gut community. Our results uncover a previously unrecognized enzymatic activity that can give rise to host-modulatory compounds and suggests a potential direct mechanism by which gut microbiota can influence host physiology, including behavior.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Anderson GM. Quantitation of tryptophan metabolites in rat feces by thin-layer chromatography. Journal of chromatography. 1975;105:323–328. - PubMed
-
- Bell SM. Treatment with gentamicin monitored by serum antibiotic assay. The Medical journal of Australia. 1976;2:481–484. - PubMed
-
- Bhattacharjee MK, Snell EE. Pyridoxal 5'-phosphate-dependent histidine decarboxylase. Mechanism of inactivation by alpha-fluoromethylhistidine. The Journal of biological chemistry. 1990;265:6664–6668. - PubMed
-
- Bianchetti MG, Minder I, Beretta-Piccoli C, Meier A, Weidmann P. Effects of tyramine on blood pressure and plasma catecholamines in normal and hypertensive subjects. Klinische Wochenschrift. 1982;60:465–470. - PubMed
-
- Bischoff SC, Mailer R, Pabst O, Weier G, Sedlik W, Li Z, Chen JJ, Murphy DL, Gershon MD. Role of serotonin in intestinal inflammation: knockout of serotonin reuptake transporter exacerbates 2,4,6-trinitrobenzene sulfonic acid colitis in mice. American journal of physiology Gastrointestinal and liver physiology. 2009;296:G685–G695. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
- DP2 OD007290/OD/NIH HHS/United States
- GM081879/GM/NIGMS NIH HHS/United States
- T32 EB009383/EB/NIBIB NIH HHS/United States
- K08 DK100638/DK/NIDDK NIH HHS/United States
- OD009180/OD/NIH HHS/United States
- DP5 OD009180/OD/NIH HHS/United States
- K08DK100638/DK/NIDDK NIH HHS/United States
- T32 GM064337/GM/NIGMS NIH HHS/United States
- P30 DK084567/DK/NIDDK NIH HHS/United States
- T32 GM008284/GM/NIGMS NIH HHS/United States
- T32 GM067547/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P50 GM081879/GM/NIGMS NIH HHS/United States
- OD007290/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
