

Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen
- PMID: 25263220
- PMCID: PMC4190086
- DOI: 10.1016/j.chom.2014.08.017
Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen
Abstract
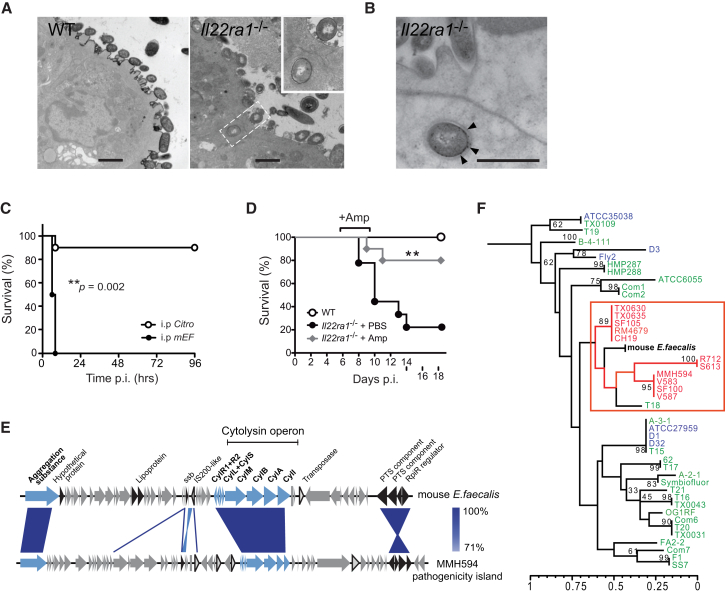
Our intestinal microbiota harbors a diverse microbial community, often containing opportunistic bacteria with virulence potential. However, mutualistic host-microbial interactions prevent disease by opportunistic pathogens through poorly understood mechanisms. We show that the epithelial interleukin-22 receptor IL-22RA1 protects against lethal Citrobacter rodentium infection and chemical-induced colitis by promoting colonization resistance against an intestinal opportunistic bacterium, Enterococcus faecalis. Susceptibility of Il22ra1(-/-) mice to C. rodentium was associated with preferential expansion and epithelial translocation of pathogenic E. faecalis during severe microbial dysbiosis and was ameloriated with antibiotics active against E. faecalis. RNA sequencing analyses of primary colonic organoids showed that IL-22RA1 signaling promotes intestinal fucosylation via induction of the fucosyltransferase Fut2. Additionally, administration of fucosylated oligosaccharides to C. rodentium-challenged Il22ra1(-/-) mice attenuated infection and promoted E. faecalis colonization resistance by restoring the diversity of anaerobic commensal symbionts. These results support a model whereby IL-22RA1 enhances host-microbiota mutualism to limit detrimental overcolonization by opportunistic pathogens.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures







Comment in
-
A delicate balance: maintaining mutualism to prevent disease.Cell Host Microbe. 2014 Oct 8;16(4):425-7. doi: 10.1016/j.chom.2014.09.019. Cell Host Microbe. 2014. PMID: 25299326 Free PMC article.
-
Tolerance: AIREs and graces.Nat Rev Immunol. 2014 Nov;14(11):714-5. doi: 10.1038/nri3762. Epub 2014 Oct 17. Nat Rev Immunol. 2014. PMID: 25324122 No abstract available.
-
Microbiota: support your friends to resist your enemies.Nat Rev Immunol. 2014 Nov;14(11):715. doi: 10.1038/nri3765. Nat Rev Immunol. 2014. PMID: 25341430 No abstract available.
References
-
- Becker D.J., Lowe J.B. Fucose: biosynthesis and biological function in mammals. Glycobiology. 2003;13:41R–53R. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
