

BS69/ZMYND11 reads and connects histone H3.3 lysine 36 trimethylation-decorated chromatin to regulated pre-mRNA processing
- PMID: 25263594
- PMCID: PMC4363072
- DOI: 10.1016/j.molcel.2014.08.022
BS69/ZMYND11 reads and connects histone H3.3 lysine 36 trimethylation-decorated chromatin to regulated pre-mRNA processing
Abstract
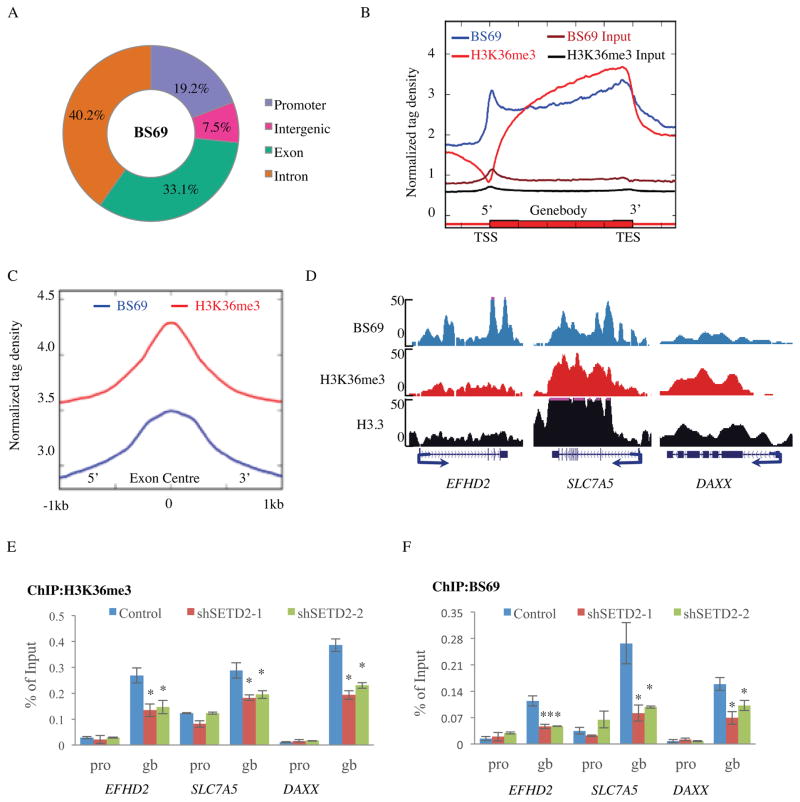
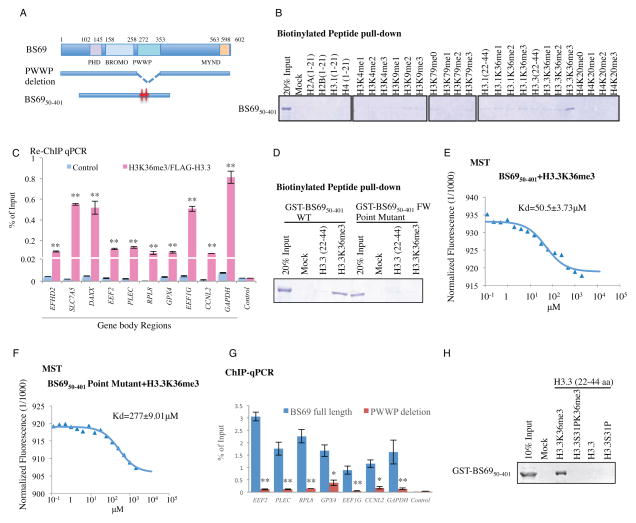
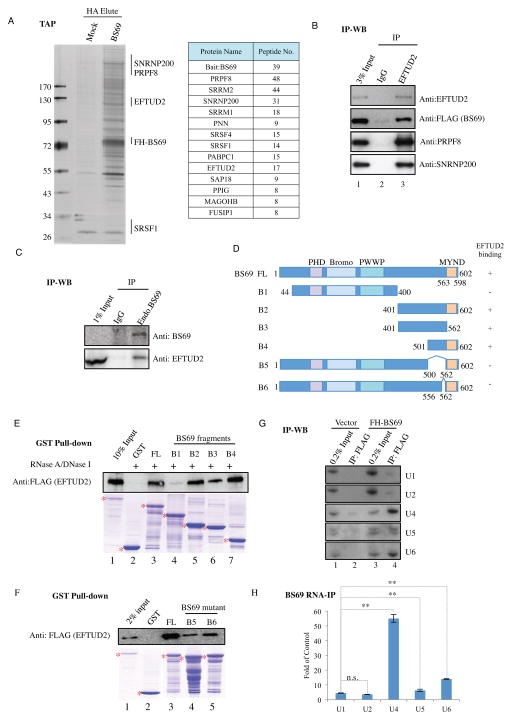
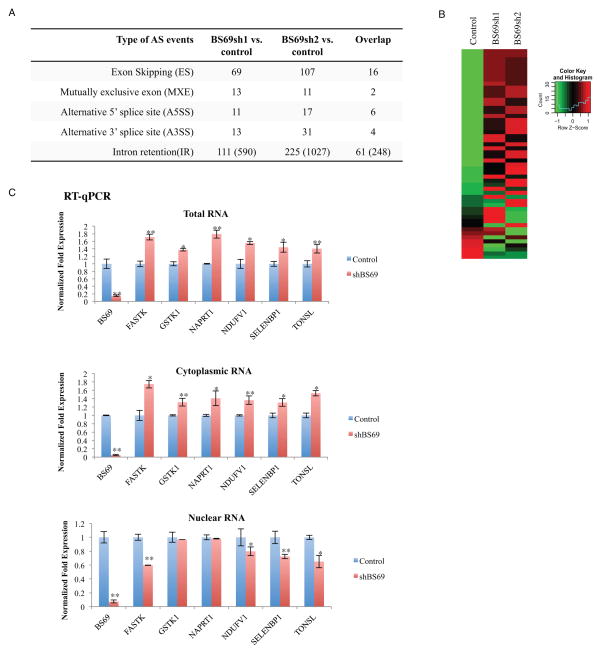
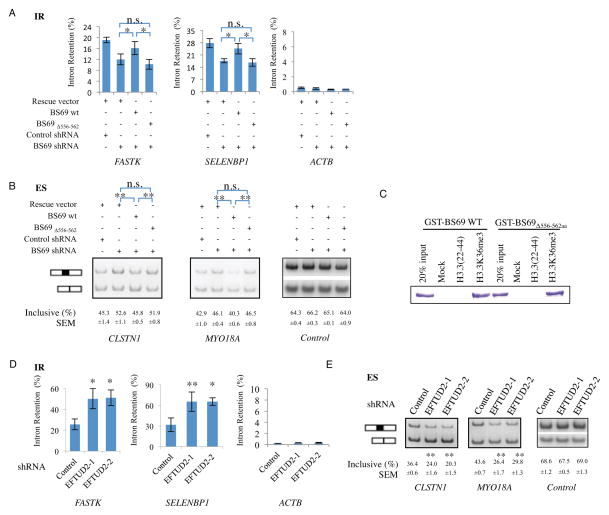
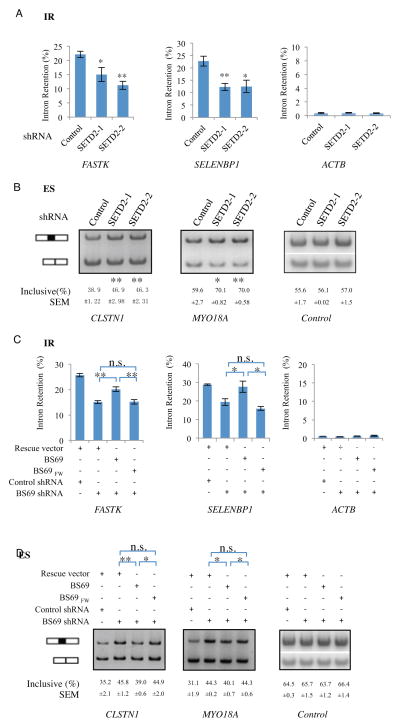
BS69 (also called ZMYND11) contains tandemly arranged PHD, BROMO, and PWWP domains, which are chromatin recognition modalities. Here, we show that BS69 selectively recognizes histone variant H3.3 lysine 36 trimethylation (H3.3K36me3) via its chromatin-binding domains. We further identify BS69 association with RNA splicing regulators, including the U5 snRNP components of the spliceosome, such as EFTUD2. Remarkably, RNA sequencing shows that BS69 mainly regulates intron retention (IR), which is the least understood RNA alternative splicing event in mammalian cells. Biochemical and genetic experiments demonstrate that BS69 promotes IR by antagonizing EFTUD2 through physical interactions. We further show that regulation of IR by BS69 also depends on its binding to H3K36me3-decorated chromatin. Taken together, our study identifies an H3.3K36me3-specific reader and a regulator of IR and reveals that BS69 connects histone H3.3K36me3 to regulated RNA splicing, providing significant, important insights into chromatin regulation of pre-mRNA processing.
Conflict of interest statement
Yang Shi is a co-fonder of Constellation Pharmaceuticals, Inc. and a member of its scientific advisory board.
Figures
References
-
- Averbeck N, Sunder S, Sample N, Wise JA, Leatherwood J. Negative control contributes to an extensive program of meiotic splicing in fission yeast. Molecular cell. 2005;18:491–498. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
