

Activation of corticotropin-releasing factor receptors in the rostral ventrolateral medulla is required for glucose-induced sympathoexcitation
- PMID: 25269482
- PMCID: PMC4233255
- DOI: 10.1152/ajpendo.00291.2014
Activation of corticotropin-releasing factor receptors in the rostral ventrolateral medulla is required for glucose-induced sympathoexcitation
Abstract
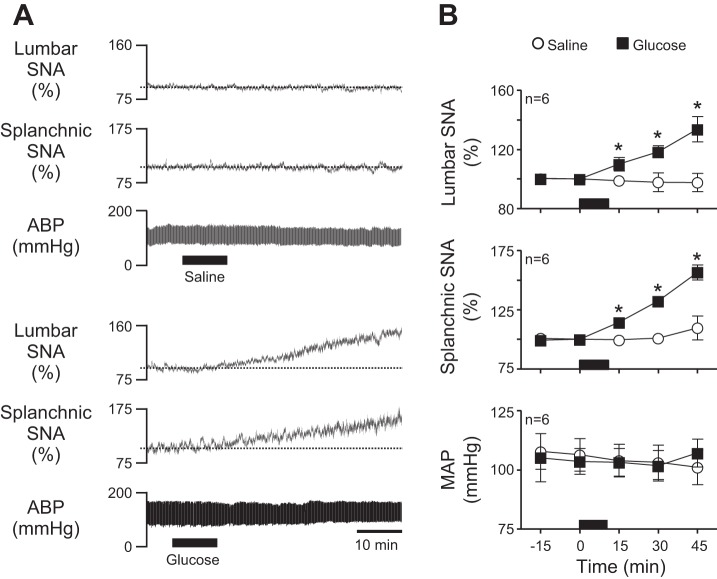
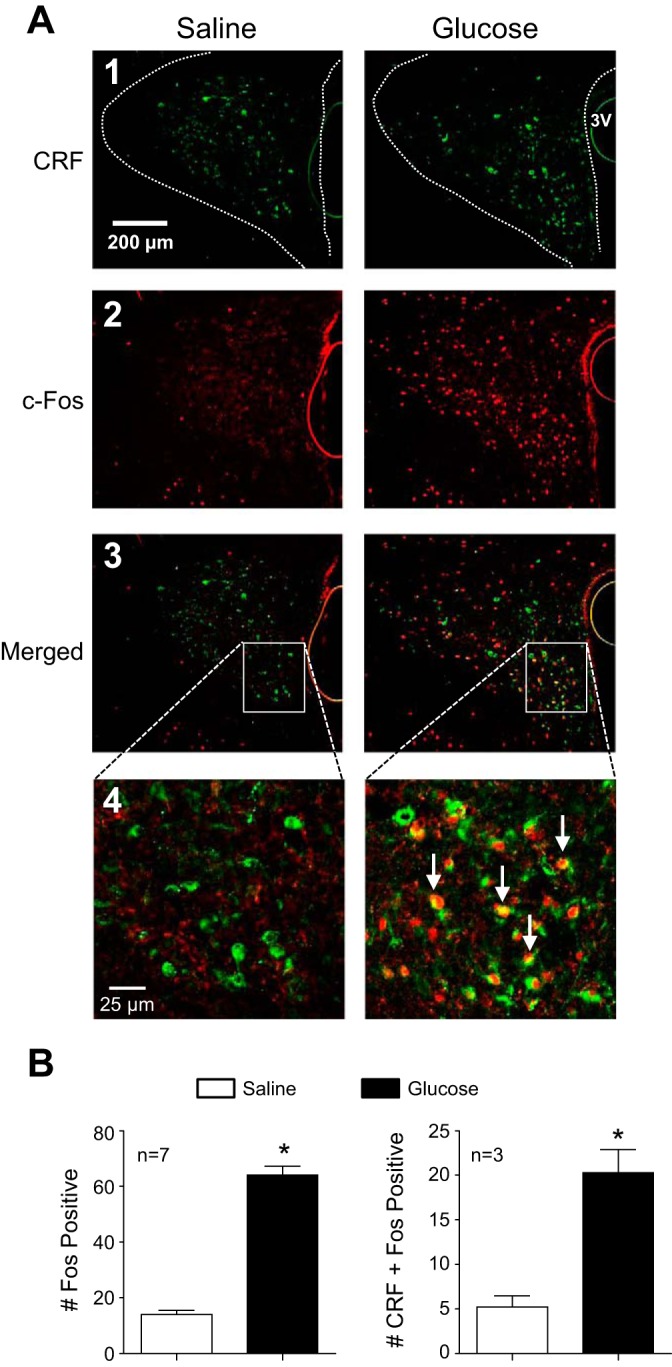
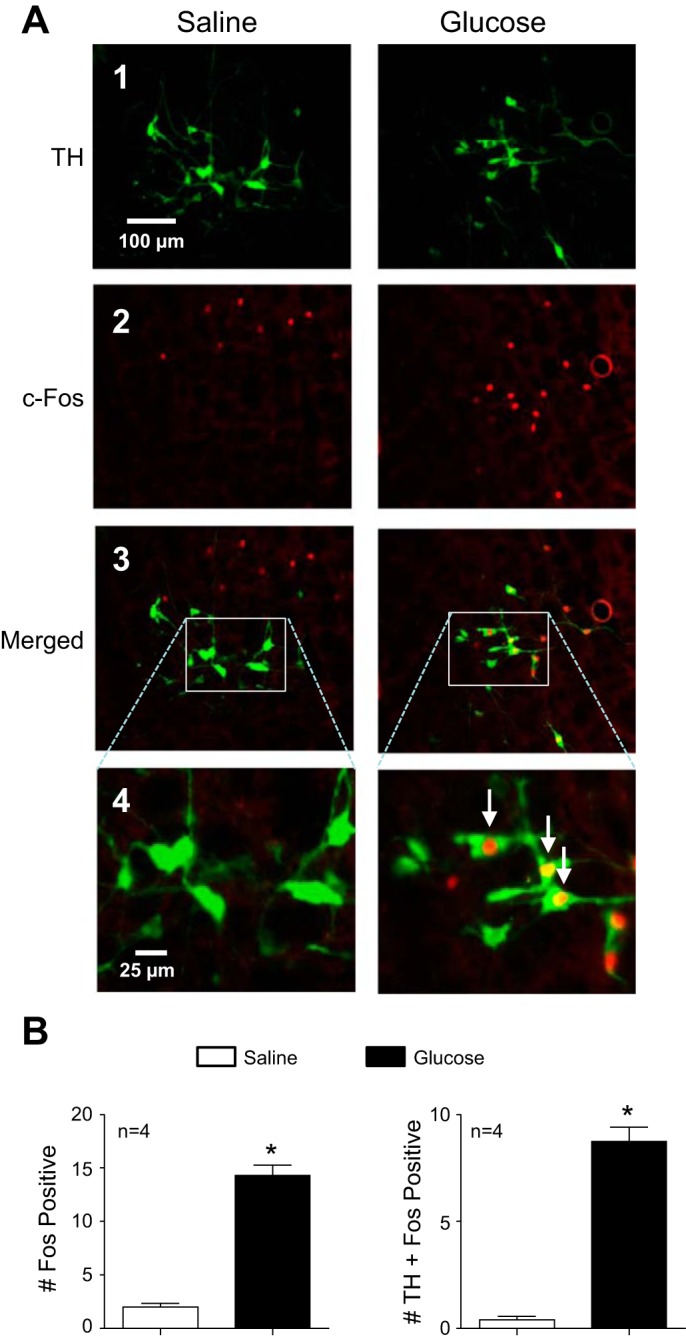
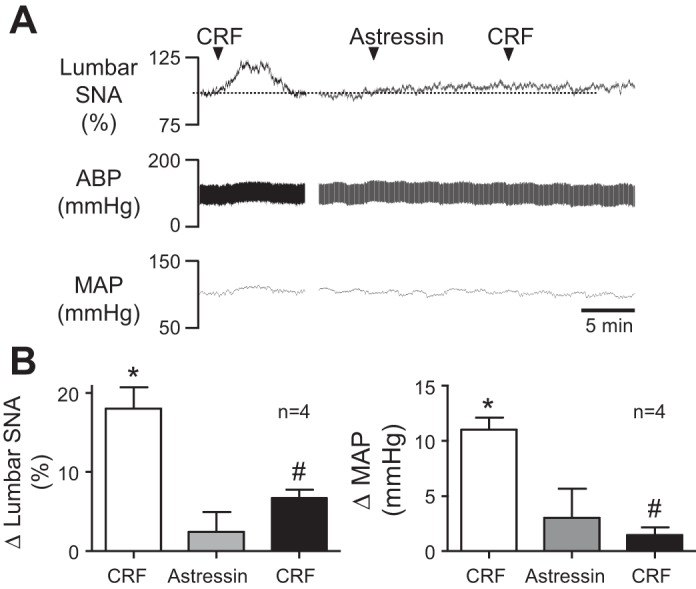
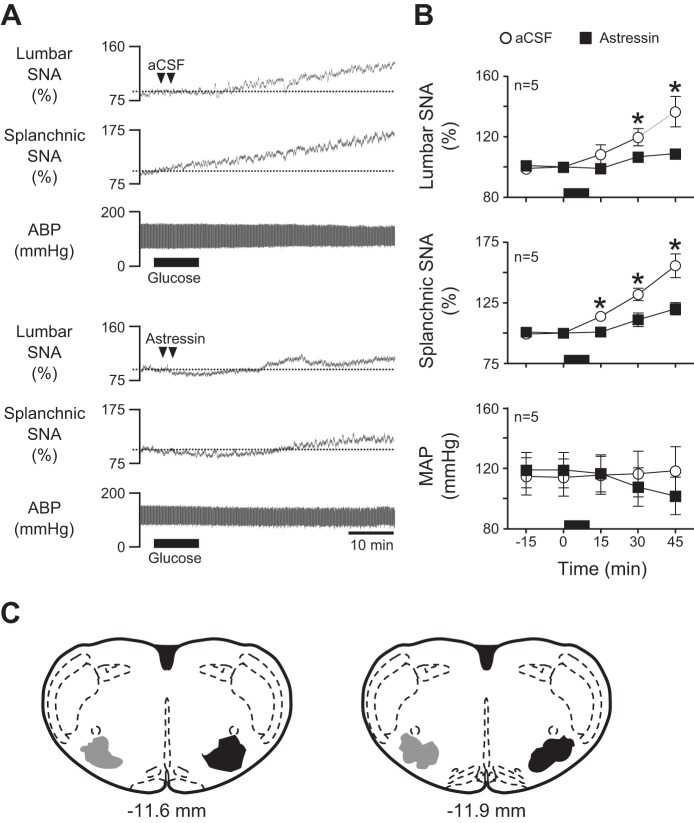
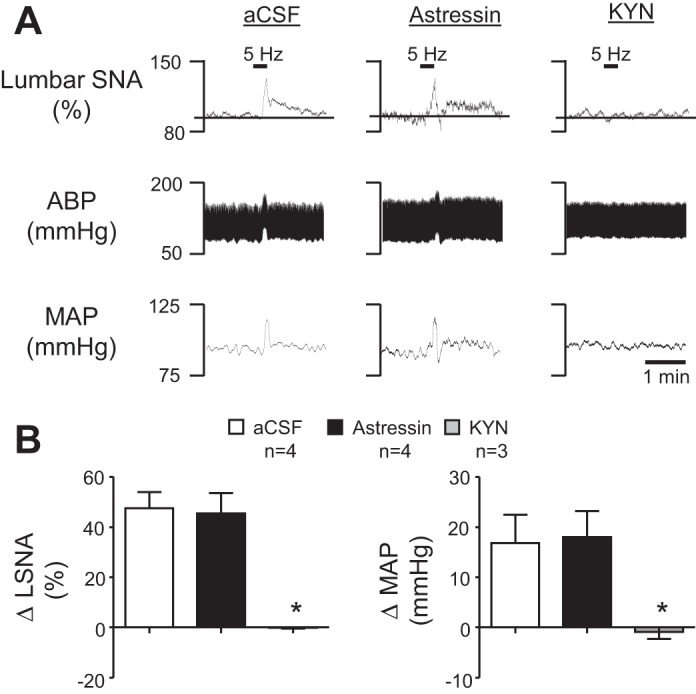
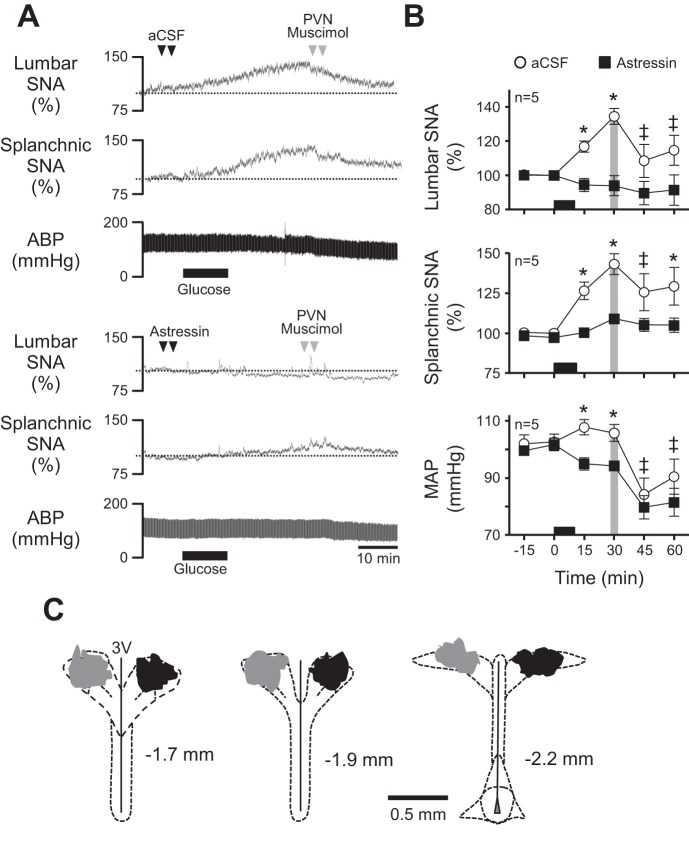
Energy expenditure is determined by metabolic rate and diet-induced thermogenesis. Normally, energy expenditure increases due to neural mechanisms that sense plasma levels of ingested nutrients/hormones and reflexively increase sympathetic nerve activity (SNA). Here, we investigated neural mechanisms of glucose-driven sympathetic activation by determining contributions of neuronal activity in the hypothalamic paraventricular nucleus (PVN) and activation of corticotropin-releasing factor (CRF) receptors in the rostral ventrolateral medulla (RVLM). Glucose was infused intravenously (150 mg/kg, 10 min) in male rats to raise plasma glucose concentration to a physiological postprandial level. In conscious rats, glucose infusion activated CRF-containing PVN neurons and TH-containing RVLM neurons, as indexed by c-Fos immunofluorescence. In α-chloralose/urethane-anesthetized rats, glucose infusion increased lumbar and splanchnic SNA, which was nearly prevented by prior RVLM injection of the CRF receptor antagonist astressin (10 pmol/50 nl). This cannot be attributed to a nonspecific effect, as sciatic afferent stimulation increased SNA and ABP equivalently in astressin- and aCSF-injected rats. Glucose-stimulated sympathoexcitation was largely reversed during inhibition of PVN neuronal activity with the GABA-A receptor agonist muscimol (100 pmol/50 nl). The effects of astressin to prevent glucose-stimulated sympathetic activation appear to be specific to interruption of PVN drive to RVLM because RVLM injection of astressin prior to glucose infusion effectively prevented SNA from rising and prevented any fall of SNA in response to acute PVN inhibition with muscimol. These findings suggest that activation of SNA, and thus energy expenditure, by glucose is initiated by activation of CRF receptors in RVLM by descending inputs from PVN.
Keywords: corticotropin releasing factor; rostral ventrolateral medulla; sympathetic nerve activity.
Copyright © 2014 the American Physiological Society.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials