

Differential genome evolution between companion symbionts in an insect-bacterial symbiosis
- PMID: 25271287
- PMCID: PMC4196230
- DOI: 10.1128/mBio.01697-14
Differential genome evolution between companion symbionts in an insect-bacterial symbiosis
Abstract
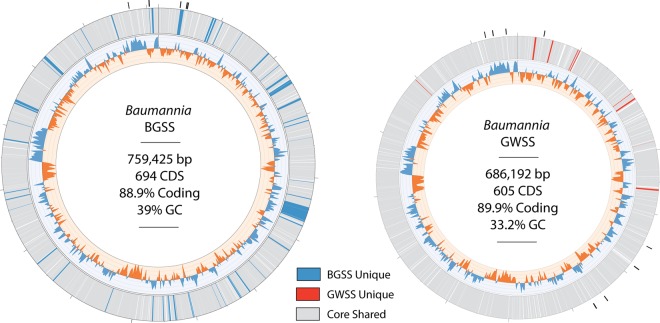
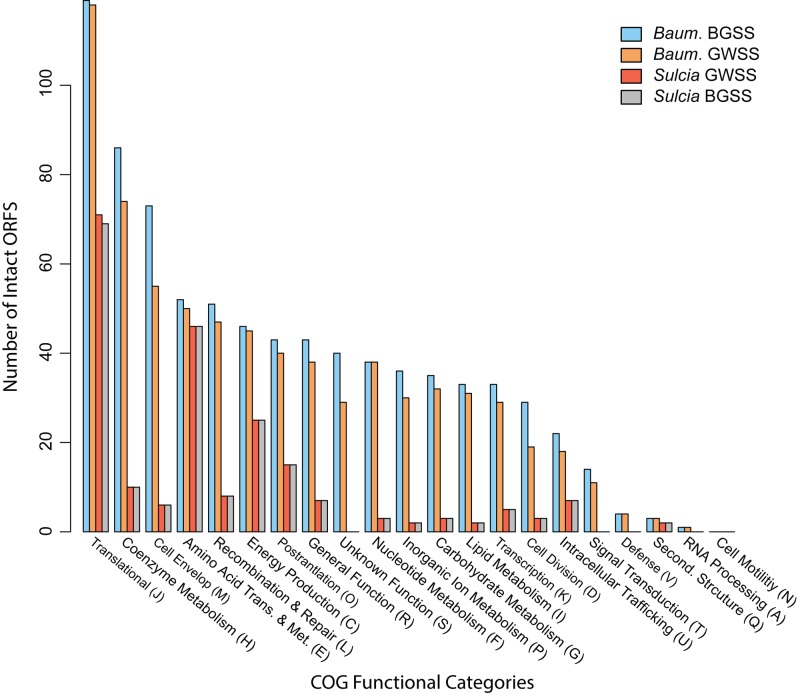
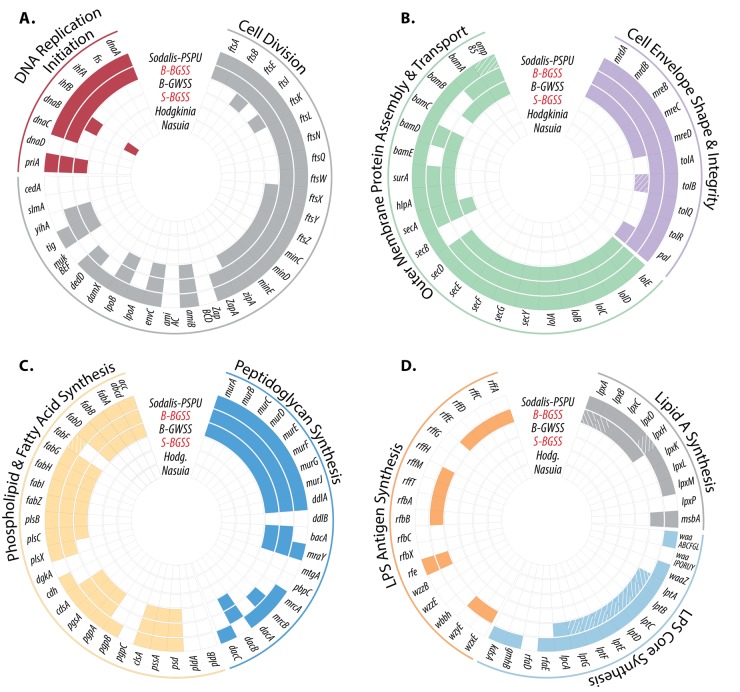
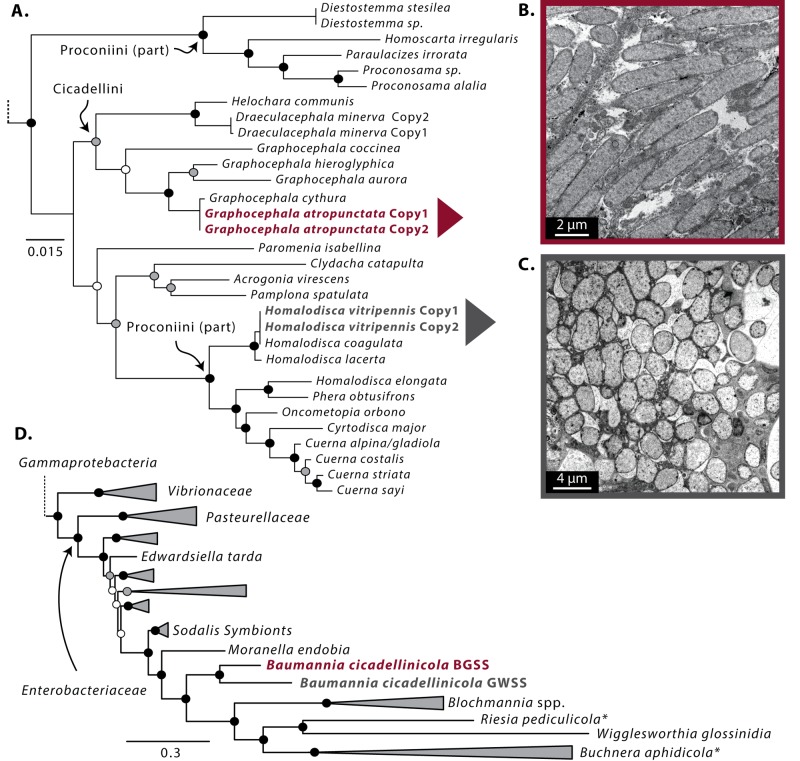
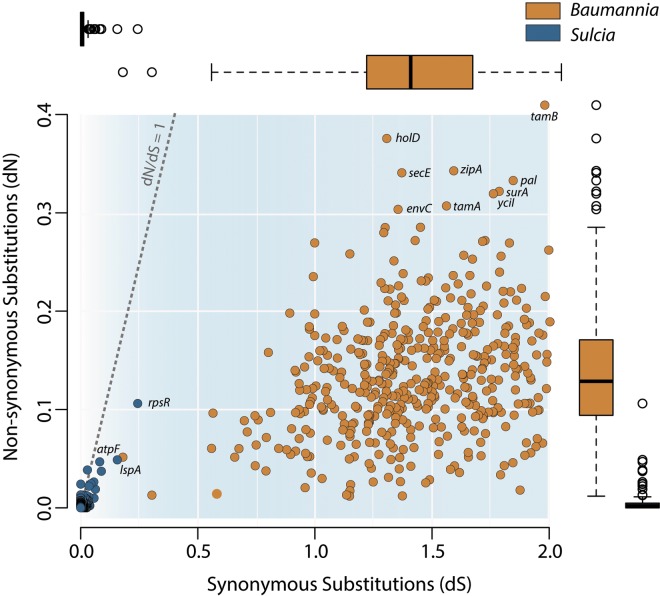
Obligate symbioses with bacteria allow insects to feed on otherwise unsuitable diets. Some symbionts have extremely reduced genomes and have lost many genes considered to be essential in other bacteria. To understand how symbiont genome degeneration proceeds, we compared the genomes of symbionts in two leafhopper species, Homalodisca vitripennis (glassy-winged sharpshooter [GWSS]) and Graphocephala atropunctata (blue-green sharpshooter [BGSS]) (Hemiptera: Cicadellidae). Each host species is associated with the anciently acquired "Candidatus Sulcia muelleri" (Bacteroidetes) and the more recently acquired "Candidatus Baumannia cicadellinicola" (Gammaproteobacteria). BGSS "Ca. Baumannia" retains 89 genes that are absent from GWSS "Ca. Baumannia"; these underlie central cellular functions, including cell envelope biogenesis, cellular replication, and stress response. In contrast, "Ca. Sulcia" strains differ by only a few genes. Although GWSS "Ca. Baumannia" cells are spherical or pleomorphic (a convergent trait of obligate symbionts), electron microscopy reveals that BGSS "Ca. Baumannia" maintains a rod shape, possibly due to its retention of genes involved in cell envelope biogenesis and integrity. Phylogenomic results suggest that "Ca. Baumannia" is derived from the clade consisting of Sodalis and relatives, a group that has evolved symbiotic associations with numerous insect hosts. Finally, the rates of synonymous and nonsynonymous substitutions are higher in "Ca. Baumannia" than in "Ca. Sulcia," which may be due to a lower mutation rate in the latter. Taken together, our results suggest that the two "Ca. Baumannia" genomes represent different stages of genome reduction in which many essential functions are being lost and likely compensated by hosts. "Ca. Sulcia" exhibits much greater genome stability and slower sequence evolution, although the mechanisms underlying these differences are poorly understood.
Importance: In obligate animal-bacterial symbioses, bacteria experience extreme patterns of genome evolution, including massive gene loss and rapid evolution. However, little is known about this process, particularly in systems with complementary bacterial partners. To understand whether genome evolution impacts symbiont types equally and whether lineages follow the same evolutionary path, we sequenced the genomes of two coresident symbiotic bacteria from a plant sap-feeding insect and compared them to the symbionts from a related host species. We found that the older symbiont has a highly reduced genome with low rates of mutation and gene loss. In contrast, the younger symbiont has a larger genome that exhibits higher mutation rates and varies dramatically in the retention of genes related to cell wall biogenesis, cellular replication, and stress response. We conclude that while symbiotic bacteria evolve toward tiny genomes, this process is shaped by different selection intensities that may reflect the different ages and metabolic roles of symbiont types.
Copyright © 2014 Bennett et al.
Figures
References
-
- Buchner P. 1965. Endosymbiosis of animals with plant microorganisms. John Wiley & Sons, New York, NY.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
