

A starting point for fluorescence-based single-molecule measurements in biomolecular research
- PMID: 25271426
- PMCID: PMC6271140
- DOI: 10.3390/molecules191015824
A starting point for fluorescence-based single-molecule measurements in biomolecular research
Abstract
Single-molecule fluorescence techniques are ideally suited to provide information about the structure-function-dynamics relationship of a biomolecule as static and dynamic heterogeneity can be easily detected. However, what type of single-molecule fluorescence technique is suited for which kind of biological question and what are the obstacles on the way to a successful single-molecule microscopy experiment? In this review, we provide practical insights into fluorescence-based single-molecule experiments aiming for scientists who wish to take their experiments to the single-molecule level. We especially focus on fluorescence resonance energy transfer (FRET) experiments as these are a widely employed tool for the investigation of biomolecular mechanisms. We will guide the reader through the most critical steps that determine the success and quality of diffusion-based confocal and immobilization-based total internal reflection fluorescence microscopy. We discuss the specific chemical and photophysical requirements that make fluorescent dyes suitable for single-molecule fluorescence experiments. Most importantly, we review recently emerged photoprotection systems as well as passivation and immobilization strategies that enable the observation of fluorescently labeled molecules under biocompatible conditions. Moreover, we discuss how the optical single-molecule toolkit has been extended in recent years to capture the physiological complexity of a cell making it even more relevant for biological research.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
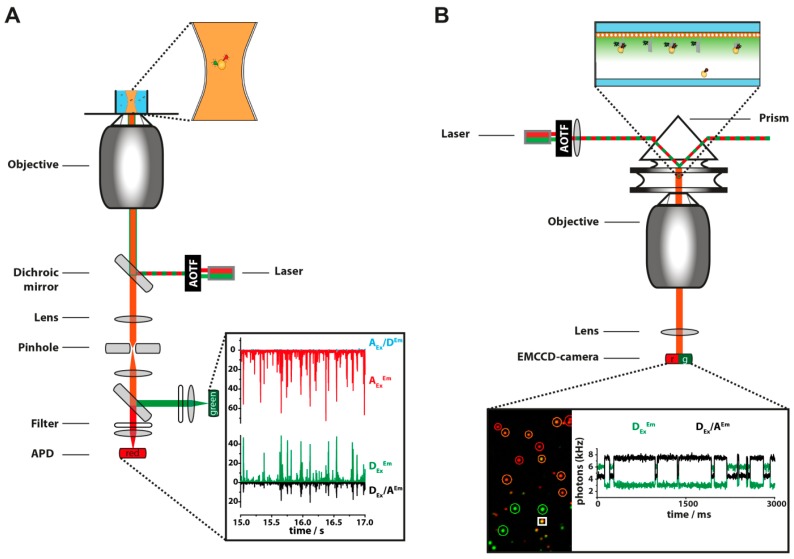
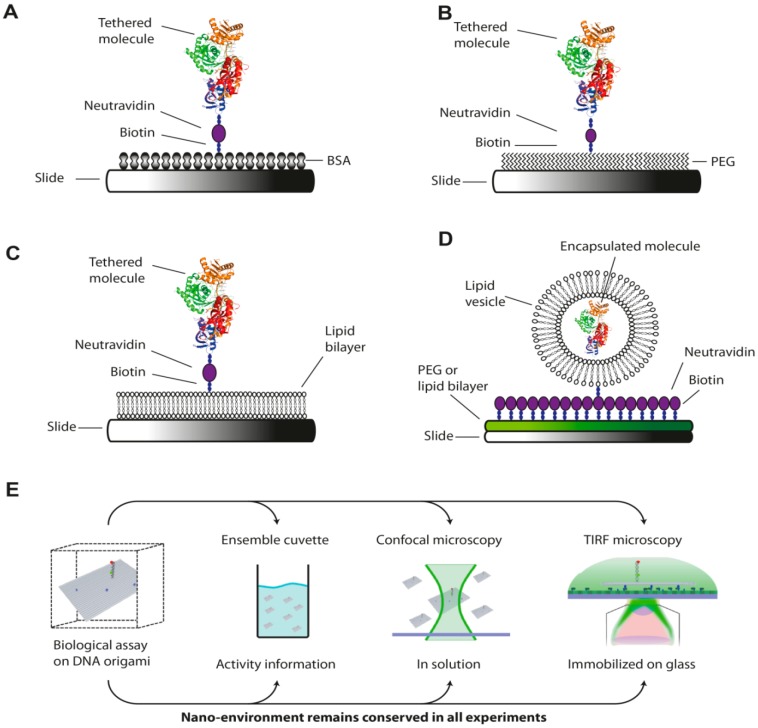
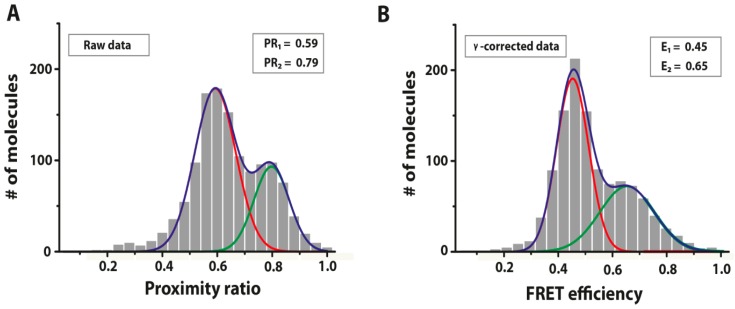
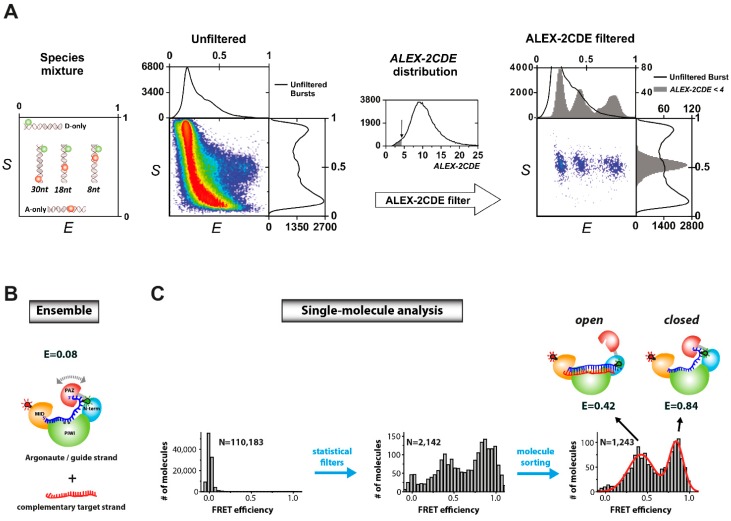
References
-
- Funatsu T., Harada Y., Tokunaga M., Saito K., Yanagida T. Imaging of single fluorescent molecules and individual atp turnovers by single myosin molecules in aqueous solution. Nature. 1995;374:555–559. - PubMed
-
- Wang Q., Moerner W.E. Single-molecule motions enable direct visualization of biomolecular interactions in solution. Nat. Methods. 2014;11:555–558. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
