

Structural insights into RNA polymerase recognition and essential function of Myxococcus xanthus CdnL
- PMID: 25272012
- PMCID: PMC4182748
- DOI: 10.1371/journal.pone.0108946
Structural insights into RNA polymerase recognition and essential function of Myxococcus xanthus CdnL
Abstract
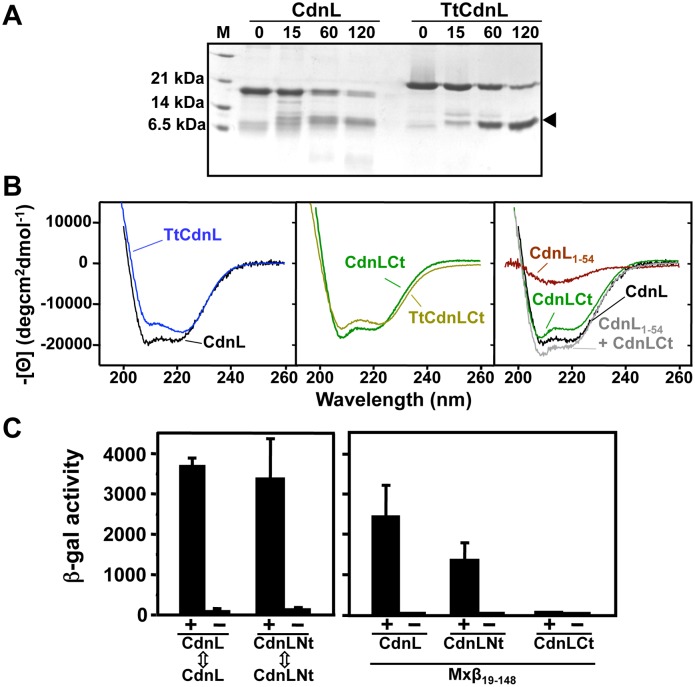
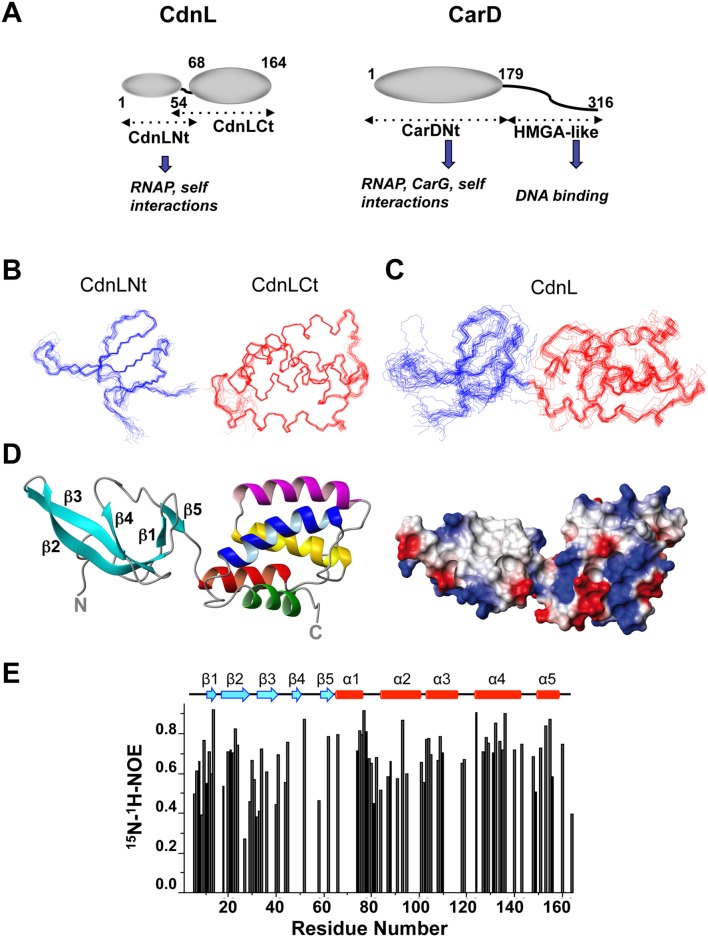
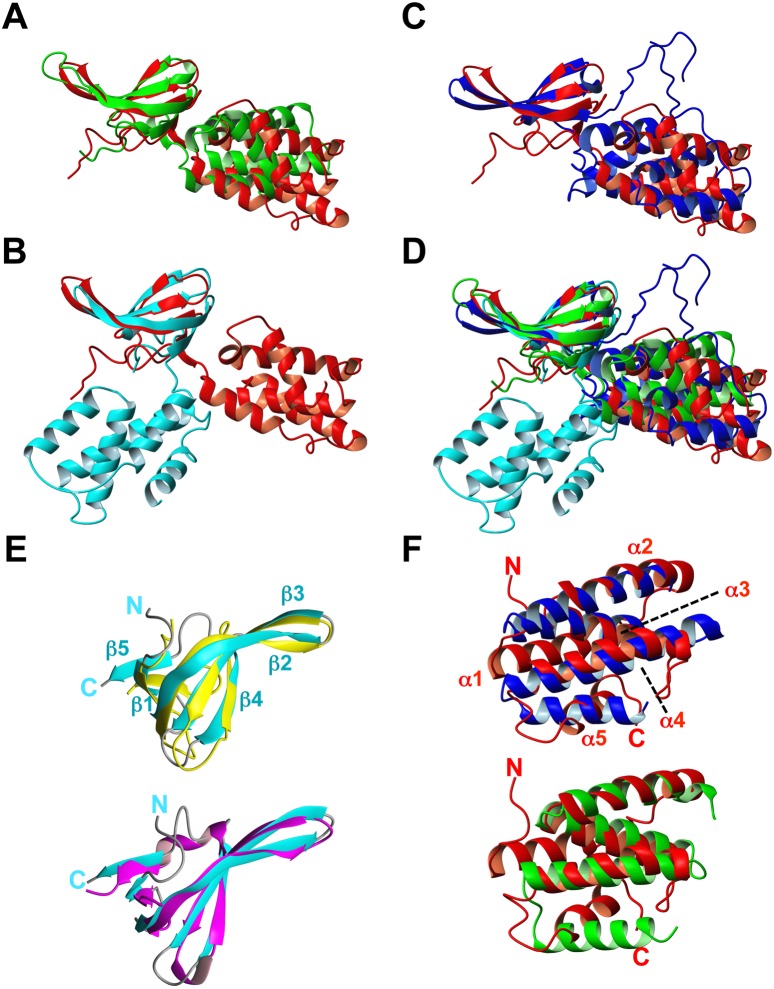
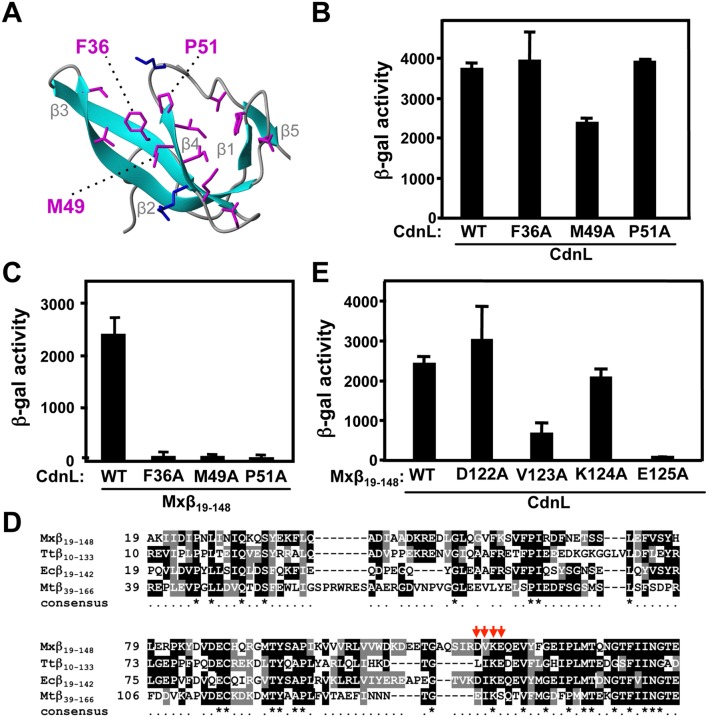
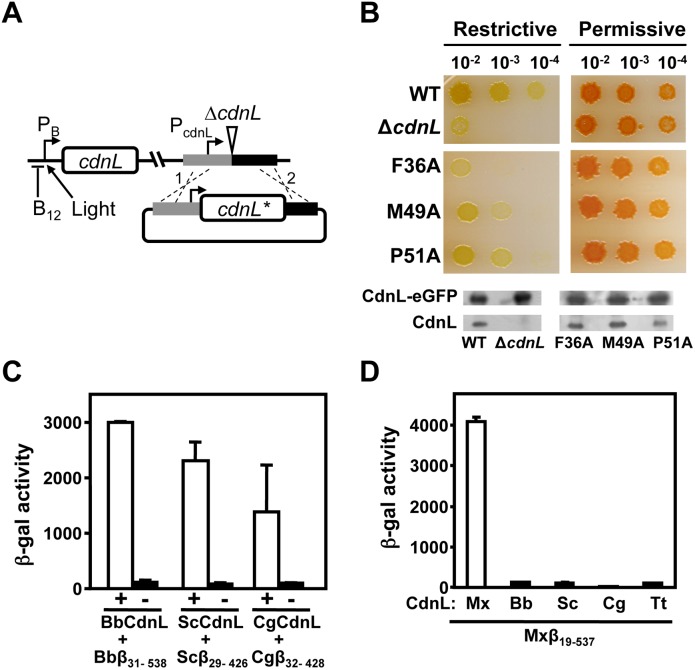
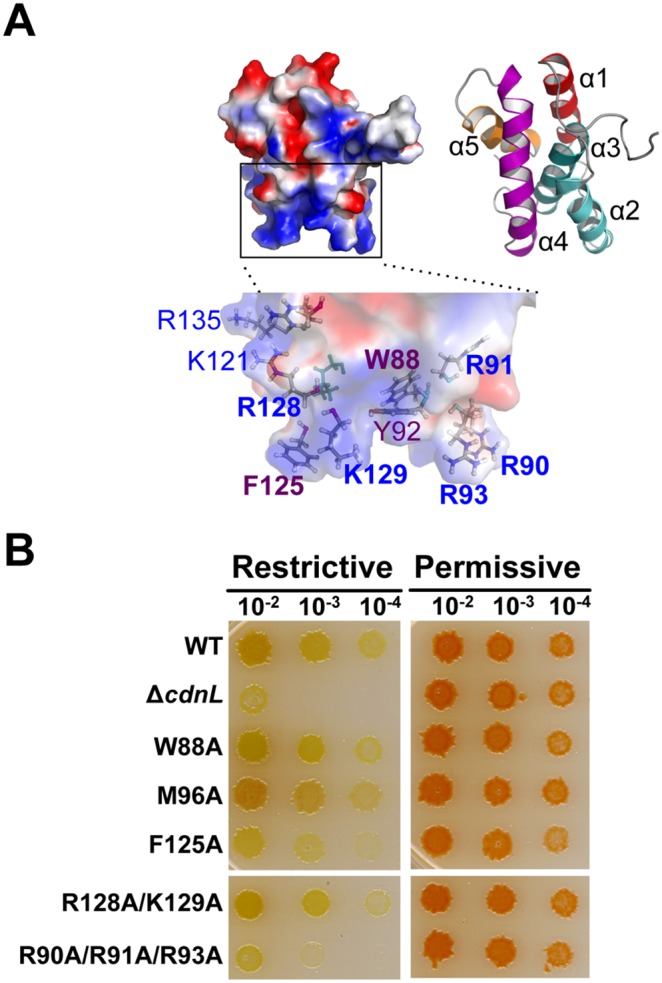
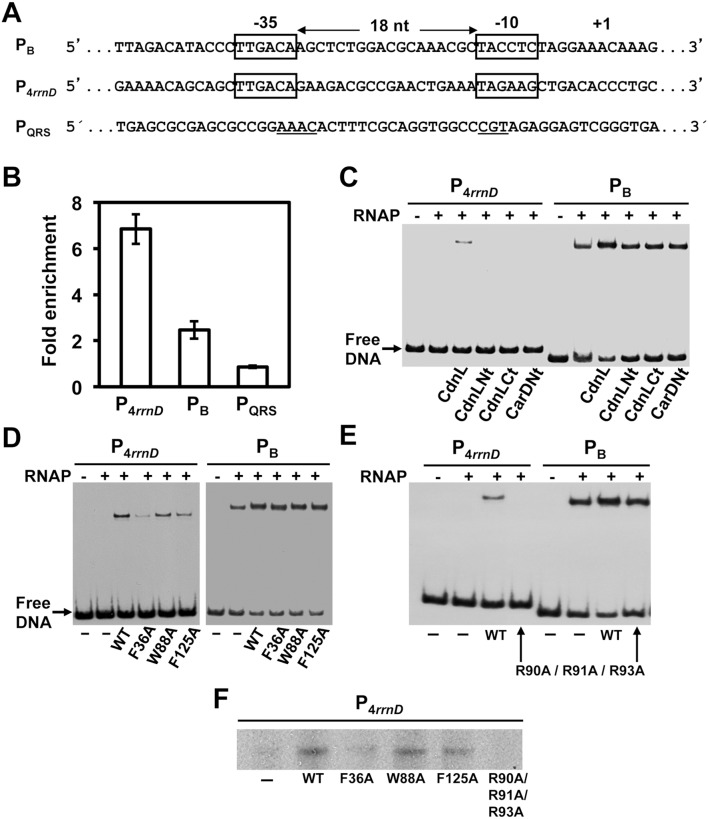
CdnL and CarD are two functionally distinct members of the CarD_CdnL_TRCF family of bacterial RNA polymerase (RNAP)-interacting proteins, which co-exist in Myxococcus xanthus. While CarD, found exclusively in myxobacteria, has been implicated in the activity of various extracytoplasmic function (ECF) σ-factors, the function and mode of action of the essential CdnL, whose homologs are widespread among bacteria, remain to be elucidated in M. xanthus. Here, we report the NMR solution structure of CdnL and present a structure-based mutational analysis of its function. An N-terminal five-stranded β-sheet Tudor-like module in the two-domain CdnL mediates binding to RNAP-β, and mutations that disrupt this interaction impair cell growth. The compact CdnL C-terminal domain consists of five α-helices folded as in some tetratricopeptide repeat-like protein-protein interaction domains, and contains a patch of solvent-exposed nonpolar and basic residues, among which a set of basic residues is shown to be crucial for CdnL function. We show that CdnL, but not its loss-of-function mutants, stabilizes formation of transcriptionally competent, open complexes by the primary σA-RNAP holoenzyme at an rRNA promoter in vitro. Consistent with this, CdnL is present at rRNA promoters in vivo. Implication of CdnL in RNAP-σA activity and of CarD in ECF-σ function in M. xanthus exemplifies how two related members within a widespread bacterial protein family have evolved to enable distinct σ-dependent promoter activity.
Conflict of interest statement
Figures
References
-
- Lee DJ, Minchin SD, Busby SJ (2012) Activating transcription in bacteria. Annu Rev Microbiol 66: 125–152. - PubMed
-
- García-Moreno D, Abellón-Ruiz J, García-Heras F, Murillo FJ, Padmanabhan S, et al. (2010) CdnL, a member of the large CarD-like family of bacterial proteins, is vital for Myxococcus xanthus and differs functionally from the global transcriptional regulator CarD. Nucleic Acids Res 38: 4586–4598. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
