

Local melatoninergic system as the protector of skin integrity
- PMID: 25272227
- PMCID: PMC4227185
- DOI: 10.3390/ijms151017705
Local melatoninergic system as the protector of skin integrity
Abstract
The human skin is not only a target for the protective actions of melatonin, but also a site of melatonin synthesis and metabolism, suggesting an important role for a local melatoninergic system in protection against ultraviolet radiation (UVR) induced damages. While melatonin exerts many effects on cell physiology and tissue homeostasis via membrane bound melatonin receptors, the strong protective effects of melatonin against the UVR-induced skin damage including DNA repair/protection seen at its high (pharmocological) concentrations indicate that these are mainly mediated through receptor-independent mechanisms or perhaps through activation of putative melatonin nuclear receptors. The destructive effects of the UVR are significantly counteracted or modulated by melatonin in the context of a complex intracutaneous melatoninergic anti-oxidative system with UVR-enhanced or UVR-independent melatonin metabolites. Therefore, endogenous intracutaneous melatonin production, together with topically-applied exogenous melatonin or metabolites would be expected to represent one of the most potent anti-oxidative defense systems against the UV-induced damage to the skin. In summary, we propose that melatonin can be exploited therapeutically as a protective agent or as a survival factor with anti-genotoxic properties or as a "guardian" of the genome and cellular integrity with clinical applications in UVR-induced pathology that includes carcinogenesis and skin aging.
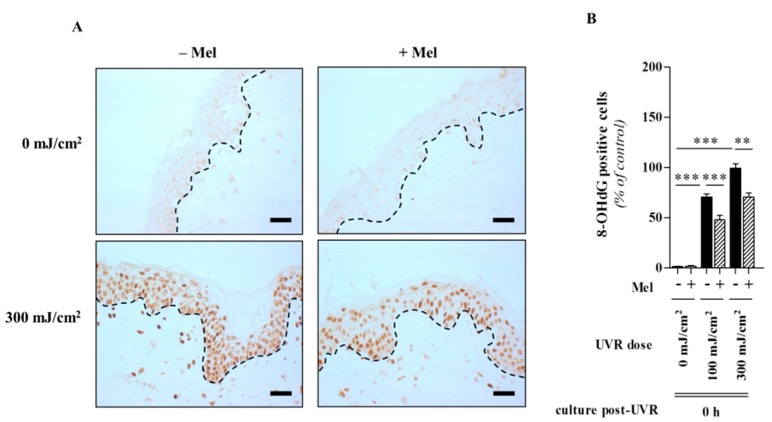
Figures
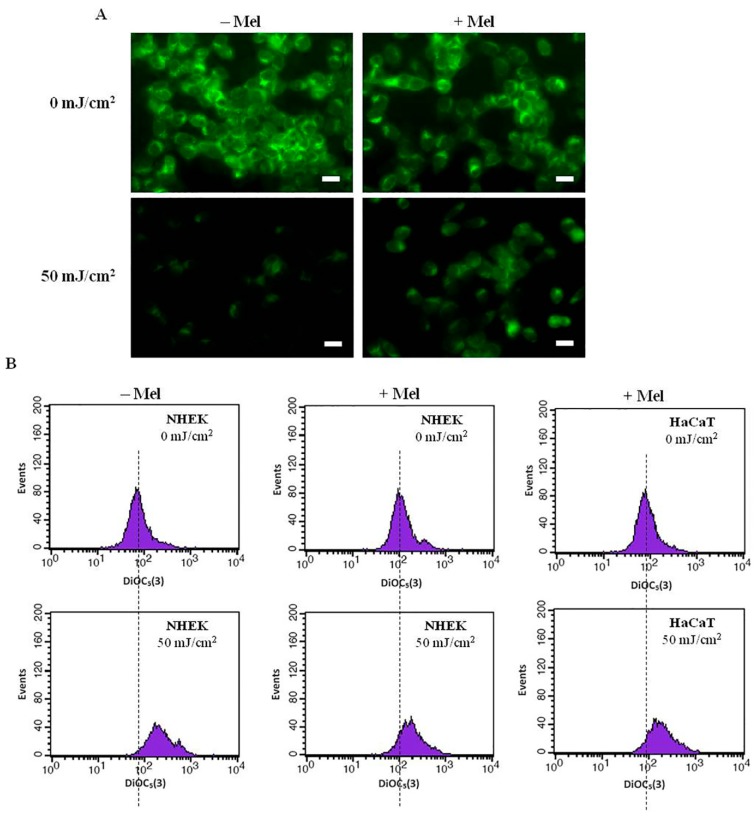
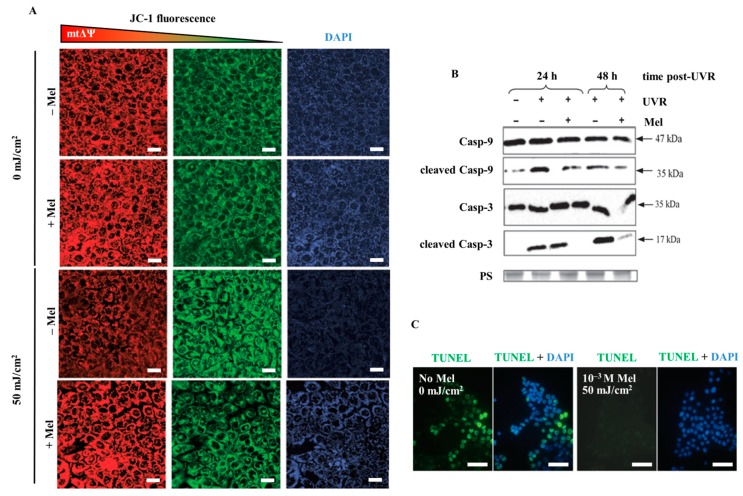
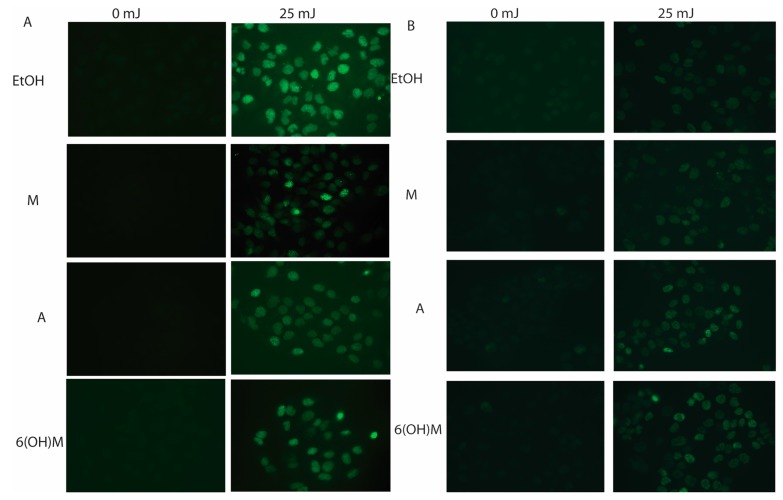
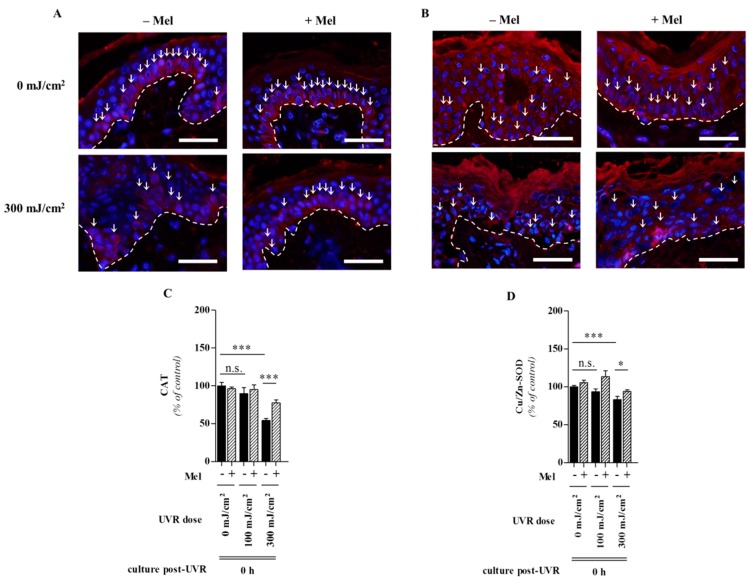
Similar articles
-
Constitutive and UV-induced metabolism of melatonin in keratinocytes and cell-free systems.FASEB J. 2006 Jul;20(9):1564-6. doi: 10.1096/fj.05-5227fje. Epub 2006 Jun 22. FASEB J. 2006. PMID: 16793870
-
Melatonin maintains mitochondrial membrane potential and attenuates activation of initiator (casp-9) and effector caspases (casp-3/casp-7) and PARP in UVR-exposed HaCaT keratinocytes.J Pineal Res. 2008 May;44(4):397-407. doi: 10.1111/j.1600-079X.2007.00542.x. Epub 2007 Dec 13. J Pineal Res. 2008. PMID: 18086147 Free PMC article.
-
Melatonin for prevention of erythema and oxidative stress in response to ultraviolet radiation.Dan Med J. 2017 Jun;64(6):B5358. Dan Med J. 2017. PMID: 28566124 Clinical Trial.
-
Melatonin in the skin: synthesis, metabolism and functions.Trends Endocrinol Metab. 2008 Jan;19(1):17-24. doi: 10.1016/j.tem.2007.10.007. Epub 2007 Dec 26. Trends Endocrinol Metab. 2008. PMID: 18155917 Review.
-
Protective Effects of Melatonin on the Skin: Future Perspectives.Int J Mol Sci. 2019 Oct 8;20(19):4948. doi: 10.3390/ijms20194948. Int J Mol Sci. 2019. PMID: 31597233 Free PMC article. Review.
Cited by
-
Ultraviolet radiation (UVR) activates central neuro-endocrine-immune system.Photodermatol Photoimmunol Photomed. 2015 May;31(3):121-3. doi: 10.1111/phpp.12165. Epub 2015 Feb 20. Photodermatol Photoimmunol Photomed. 2015. PMID: 25660238 Free PMC article. No abstract available.
-
Photoprotective Effects of Cycloheterophyllin against UVA-Induced Damage and Oxidative Stress in Human Dermal Fibroblasts.PLoS One. 2016 Sep 1;11(9):e0161767. doi: 10.1371/journal.pone.0161767. eCollection 2016. PLoS One. 2016. PMID: 27583973 Free PMC article.
-
Melatonin and the Skin: Current Progress and Perspectives for Human Health.J Invest Dermatol. 2025 Jun;145(6):1345-1360.e2. doi: 10.1016/j.jid.2024.11.012. Epub 2025 Feb 6. J Invest Dermatol. 2025. PMID: 39918482 Review.
-
The Influence of Circadian Rhythms on DNA Damage Repair in Skin Photoaging.Int J Mol Sci. 2024 Oct 11;25(20):10926. doi: 10.3390/ijms252010926. Int J Mol Sci. 2024. PMID: 39456709 Free PMC article. Review.
-
The Promising Role of Polyphenols in Skin Disorders.Molecules. 2024 Feb 15;29(4):865. doi: 10.3390/molecules29040865. Molecules. 2024. PMID: 38398617 Free PMC article. Review.
References
-
- Lerner A.B., Case J.D., Takahashi Y. Isolation of melatonin and 5-methoxyindole-3-acetic acid from bovine pineal glands. J. Biol. Chem. 1960;235:1992–1997. - PubMed
-
- Reiter R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev. 1991;12:151–180. - PubMed
-
- Friedman P.A., Kappelman A.H., Kaufman S. Partial purification and characterization of tryptophan hydroxylase from rabbit hindbrain. J. Biol. Chem. 1972;247:4165–4173. - PubMed
-
- Mockus S.M., Vrana K.E. Advances in the molecular characterization of tryptophan hydroxylase. J. Mol. Neurosci. 1998;10:163–179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
