

A biomechanical mechanism for initiating DNA packaging
- PMID: 25274732
- PMCID: PMC4231757
- DOI: 10.1093/nar/gku896
A biomechanical mechanism for initiating DNA packaging
Abstract
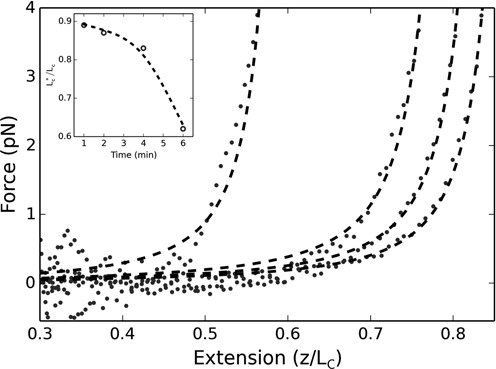
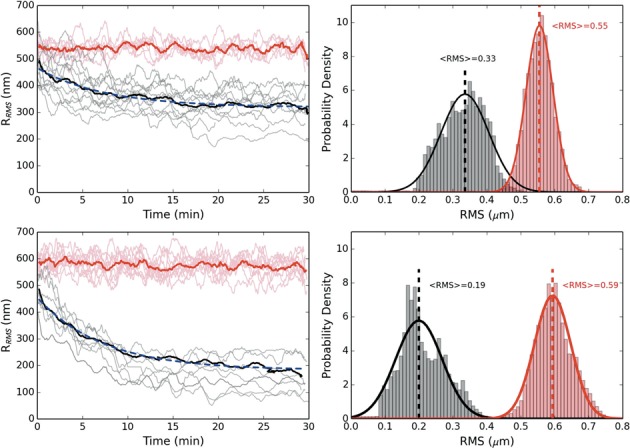
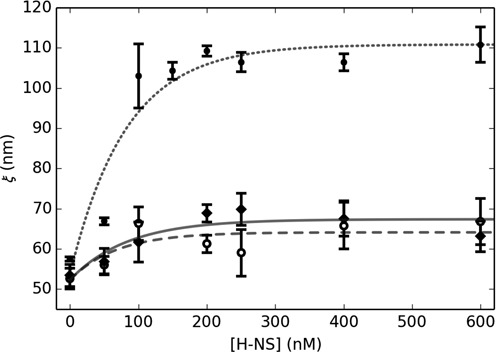
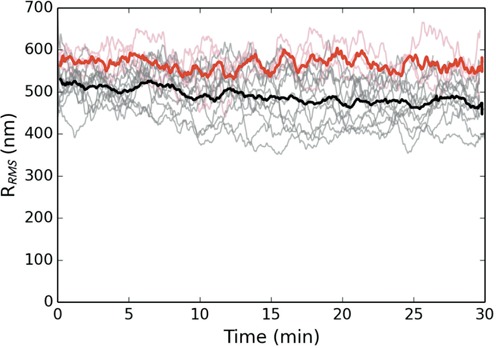
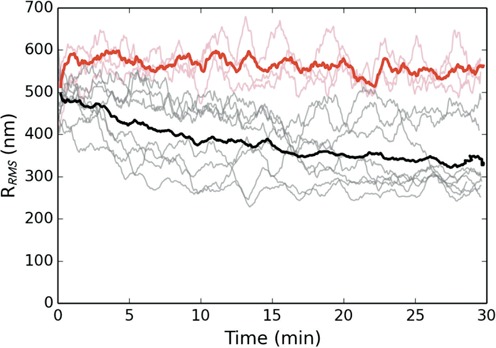
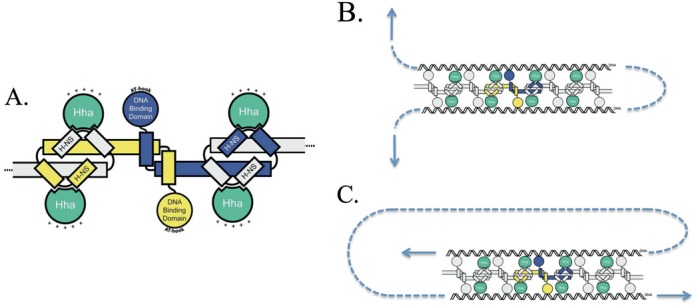
The bacterial chromosome is under varying levels of mechanical stress due to a high degree of crowding and dynamic protein-DNA interactions experienced within the nucleoid. DNA tension is difficult to measure in cells and its functional significance remains unclear although in vitro experiments have implicated a range of biomechanical phenomena. Using single-molecule tools, we have uncovered a novel protein-DNA interaction that responds to fluctuations in mechanical tension by condensing DNA. We combined tethered particle motion (TPM) and optical tweezers experiments to probe the effects of tension on DNA in the presence of the Hha/H-NS complex. The nucleoid structuring protein H-NS is a key regulator of DNA condensation and gene expression in enterobacteria and its activity in vivo is affected by the accessory factor Hha. We find that tension, induced by optical tweezers, causes the rapid compaction of DNA in the presence of the Hha/H-NS complex, but not in the presence of H-NS alone. Our results imply that H-NS requires Hha to condense bacterial DNA and that this condensation could be triggered by the level of mechanical tension experienced along different regions of the chromosome.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Ochman H., Lawrence J.G., Groisman E.A. Lateral gene transfer and the nature of bacterial innovation. Nature. 2000;405:299–304. - PubMed
-
- Gevers D., Cohan F.M., Lawrence J.G., Spratt B.G., Coenye T., Feil E.J., Stackebrandt E., Van de Peer Y., Vandamme P., Thompson F.L., et al. Opinion: re-evaluating prokaryotic species. Nat. Rev. Microbiol. 2005;3:733–739. - PubMed
-
- Jain R., Rivera M.C., Moore J.E., Lake J.A. Horizontal gene transfer in microbial genome evolution. Theor. Popul. Biol. 2002;61:489–495. - PubMed
-
- Thanbichler M., Wang S.C., Shapiro L. The bacterial nucleoid: a highly organized and dynamic structure. J. Cell. Biochem. 2005;96:506–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
