

Self-consistent determination of the spike-train power spectrum in a neural network with sparse connectivity
- PMID: 25278869
- PMCID: PMC4166962
- DOI: 10.3389/fncom.2014.00104
Self-consistent determination of the spike-train power spectrum in a neural network with sparse connectivity
Abstract
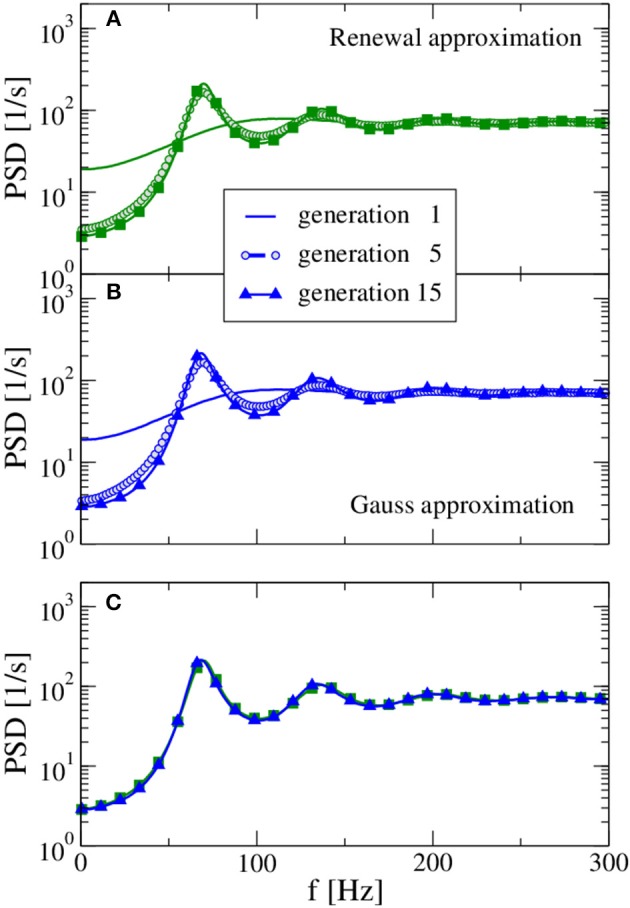
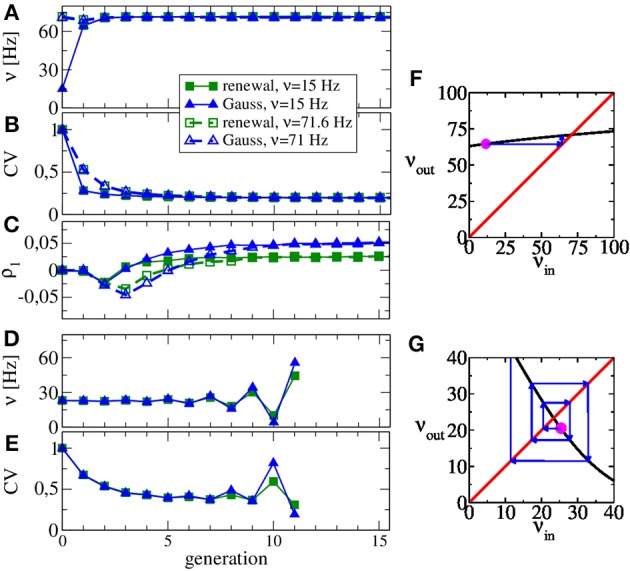
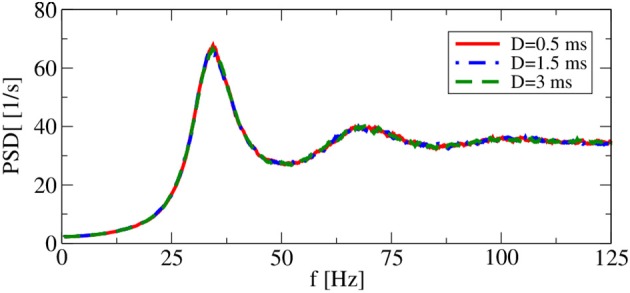
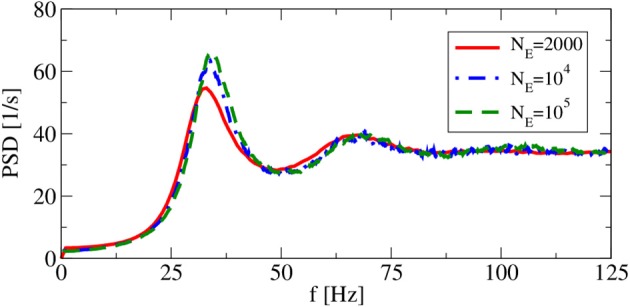
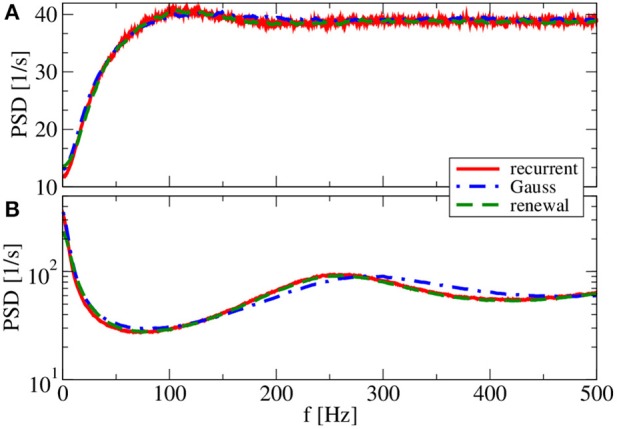
A major source of random variability in cortical networks is the quasi-random arrival of presynaptic action potentials from many other cells. In network studies as well as in the study of the response properties of single cells embedded in a network, synaptic background input is often approximated by Poissonian spike trains. However, the output statistics of the cells is in most cases far from being Poisson. This is inconsistent with the assumption of similar spike-train statistics for pre- and postsynaptic cells in a recurrent network. Here we tackle this problem for the popular class of integrate-and-fire neurons and study a self-consistent statistics of input and output spectra of neural spike trains. Instead of actually using a large network, we use an iterative scheme, in which we simulate a single neuron over several generations. In each of these generations, the neuron is stimulated with surrogate stochastic input that has a similar statistics as the output of the previous generation. For the surrogate input, we employ two distinct approximations: (i) a superposition of renewal spike trains with the same interspike interval density as observed in the previous generation and (ii) a Gaussian current with a power spectrum proportional to that observed in the previous generation. For input parameters that correspond to balanced input in the network, both the renewal and the Gaussian iteration procedure converge quickly and yield comparable results for the self-consistent spike-train power spectrum. We compare our results to large-scale simulations of a random sparsely connected network of leaky integrate-and-fire neurons (Brunel, 2000) and show that in the asynchronous regime close to a state of balanced synaptic input from the network, our iterative schemes provide an excellent approximations to the autocorrelation of spike trains in the recurrent network.
Keywords: neural noise; non-Poissonian spiking; recurrent neural networks; spike-train power spectrum; spike-train statistics.
Figures

References
-
- Bahar S., Kantelhardt J. W., Neiman A., Rego H. H. A., Russell D. F., Wilkens L., et al. (2001). Long-range temporal anti-correlations in paddlefish electroreceptors. Europhys. Lett. 56, 454–460 10.1209/epl/i2001-00540-7 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources