

In search of a recognition memory engram
- PMID: 25280908
- PMCID: PMC4382520
- DOI: 10.1016/j.neubiorev.2014.09.016
In search of a recognition memory engram
Abstract
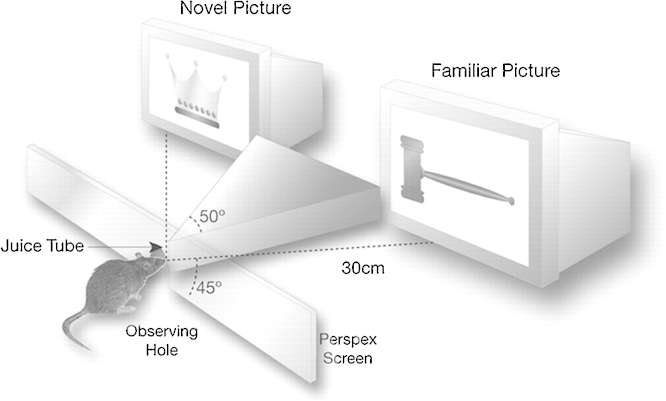
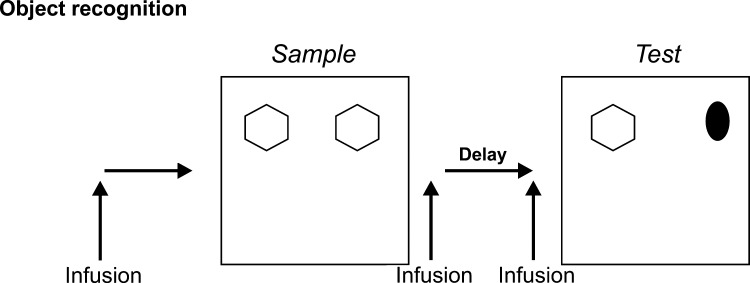
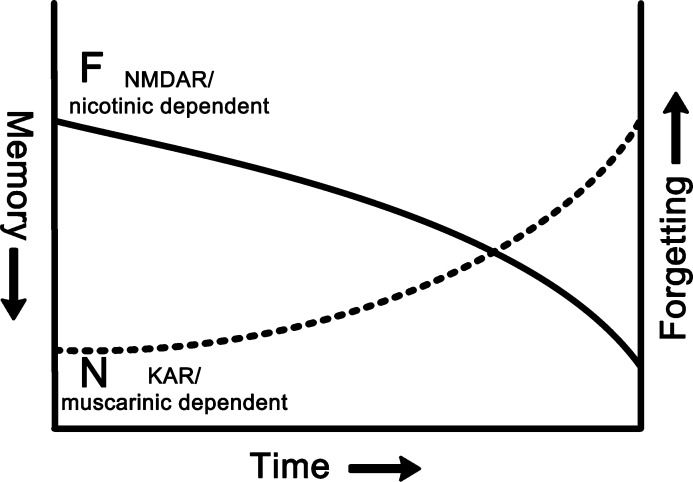
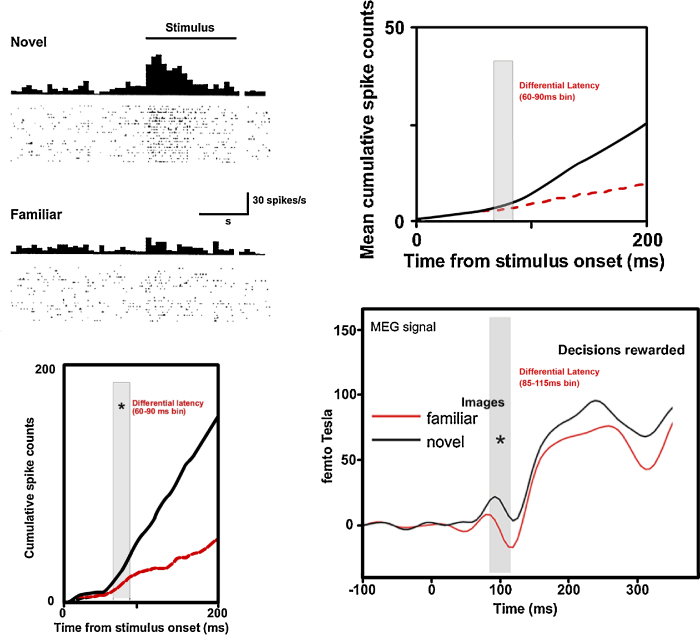
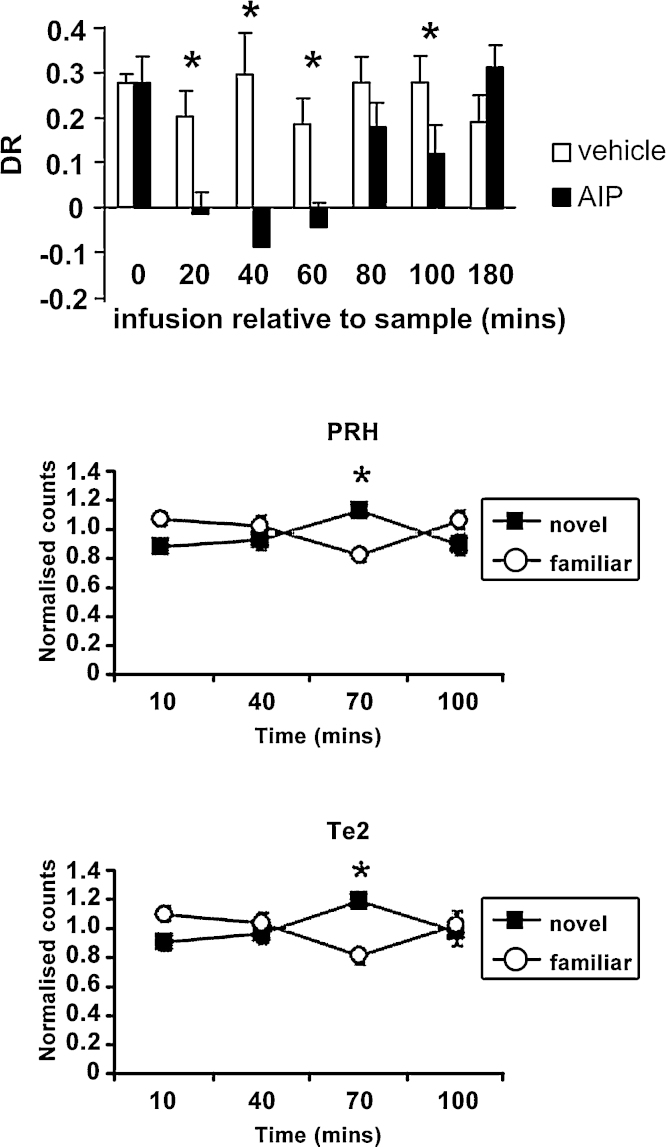
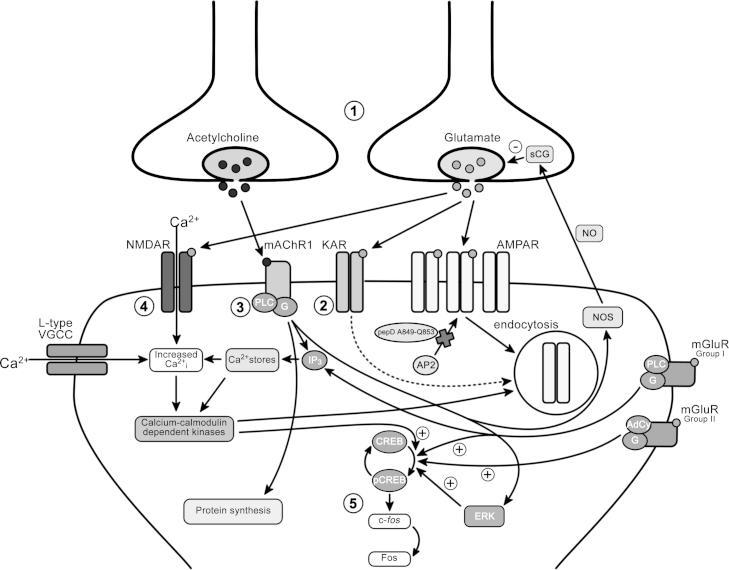
A large body of data from human and animal studies using psychological, recording, imaging, and lesion techniques indicates that recognition memory involves at least two separable processes: familiarity discrimination and recollection. Familiarity discrimination for individual visual stimuli seems to be effected by a system centred on the perirhinal cortex of the temporal lobe. The fundamental change that encodes prior occurrence within the perirhinal cortex is a reduction in the responses of neurones when a stimulus is repeated. Neuronal network modelling indicates that a system based on such a change in responsiveness is potentially highly efficient in information theoretic terms. A review is given of findings indicating that perirhinal cortex acts as a storage site for recognition memory of objects and that such storage depends upon processes producing synaptic weakening.
Keywords: Familiarity; Imprinting; LTD; LTP; Perirhinal cortex.
Copyright © 2014 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Abe H., Ishida Y., Iwasaki T. Perirhinal N-methyl-d-aspartate and muscarinic systems participate in object recognition in rats. Neurosci. Lett. 2004;356:191–194. - PubMed
-
- Abe H., Iwasaki T. NMDA and muscarinic blockade in the perirhinal cortex impairs object discrimination in rats. Neuroreport. 2001;12:3375–3379. - PubMed
-
- Aggleton J.P., Brown M.W., Albasser M.M. Contrasting brain activity patterns for item recognition memory and associative recognition memory: insights from immediate-early gene functional imaging. Neuropsychologia. 2012;50:3141–3155. - PubMed
-
- Albasser M., Davies M., Futter J., Aggleton J. Magnitude of the object recognition deficit associated with perirhinal cortex damage in rats: effects of varying the lesion extent and the duration of the sample period. Behav. Neurosci. 2009;123:115–124. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
