

Arabidopsis PHOSPHOTYROSYL PHOSPHATASE ACTIVATOR is essential for PROTEIN PHOSPHATASE 2A holoenzyme assembly and plays important roles in hormone signaling, salt stress response, and plant development
- PMID: 25281708
- PMCID: PMC4226365
- DOI: 10.1104/pp.114.250563
Arabidopsis PHOSPHOTYROSYL PHOSPHATASE ACTIVATOR is essential for PROTEIN PHOSPHATASE 2A holoenzyme assembly and plays important roles in hormone signaling, salt stress response, and plant development
Abstract
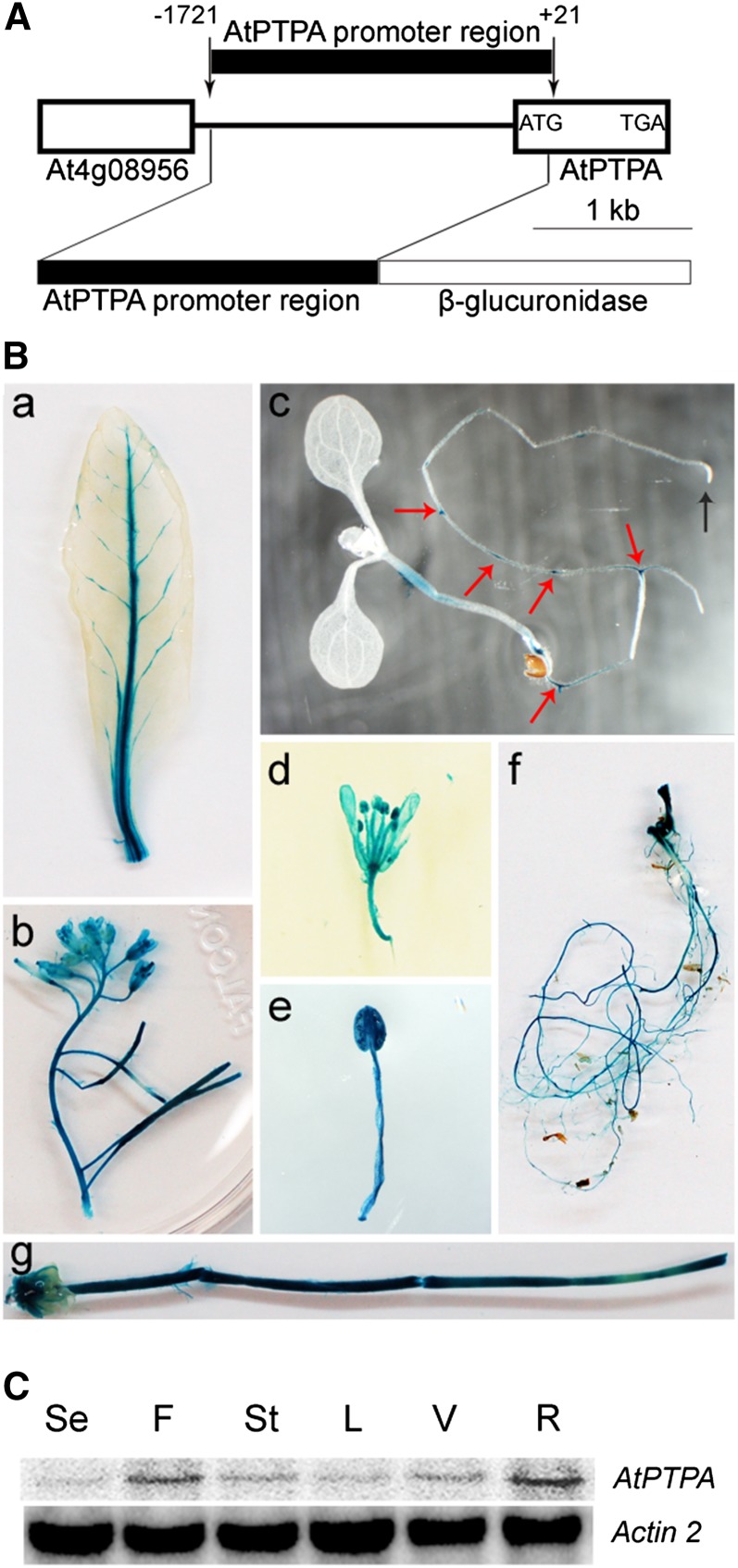
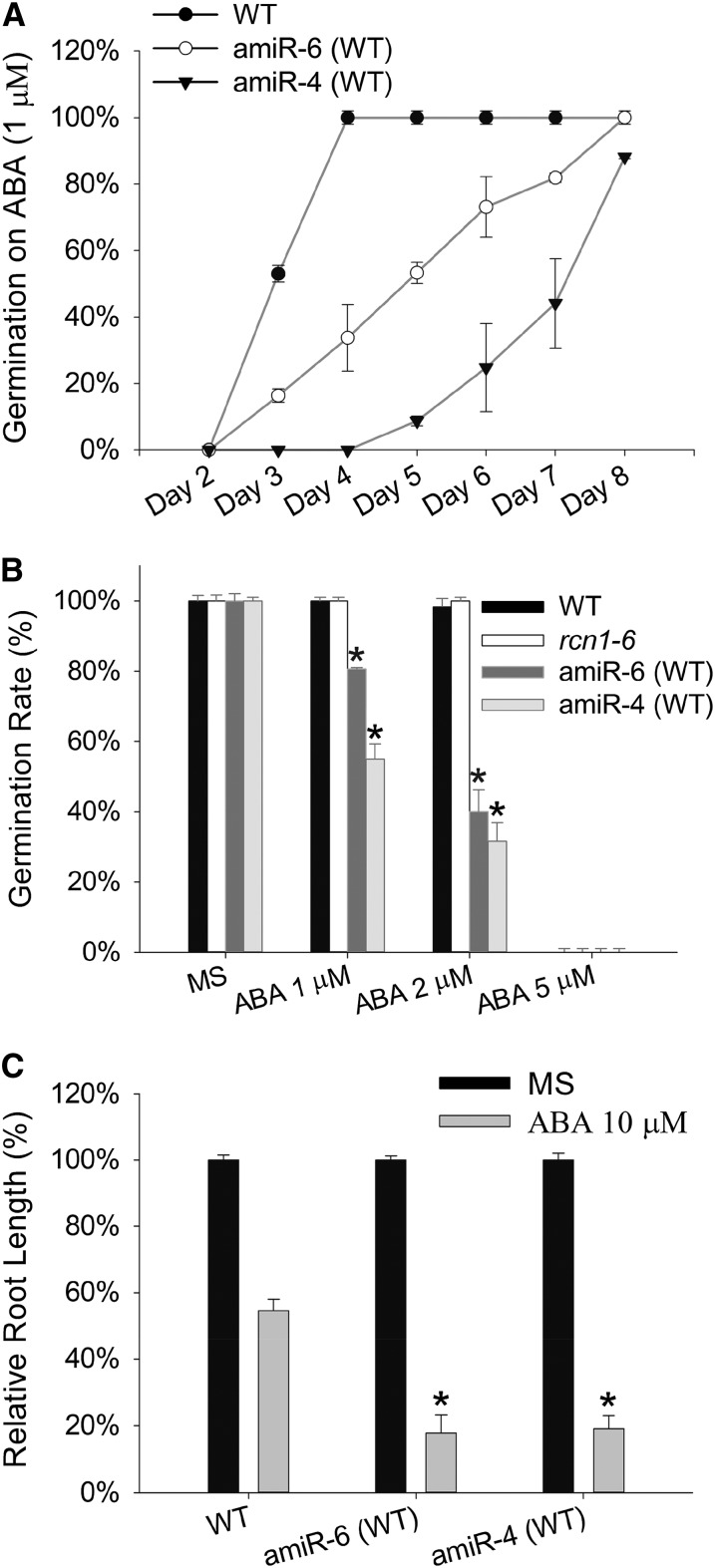
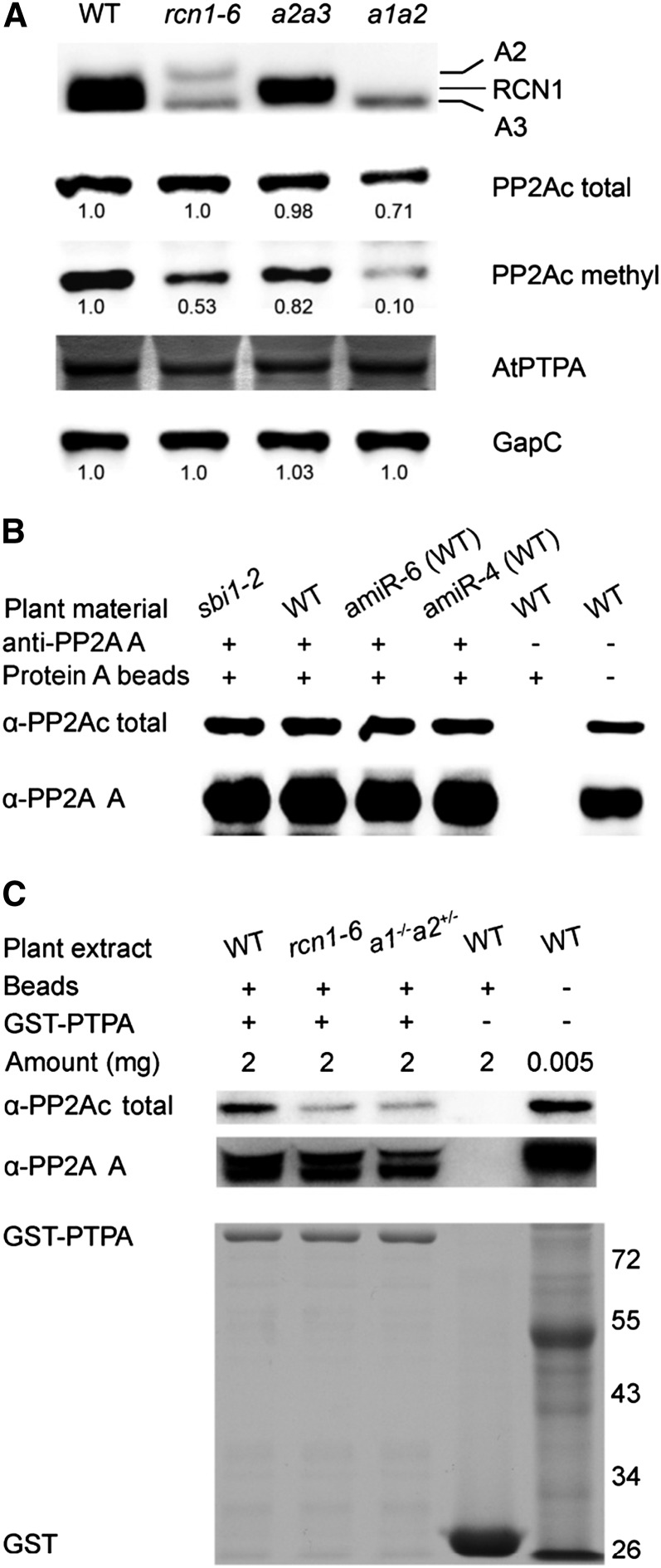
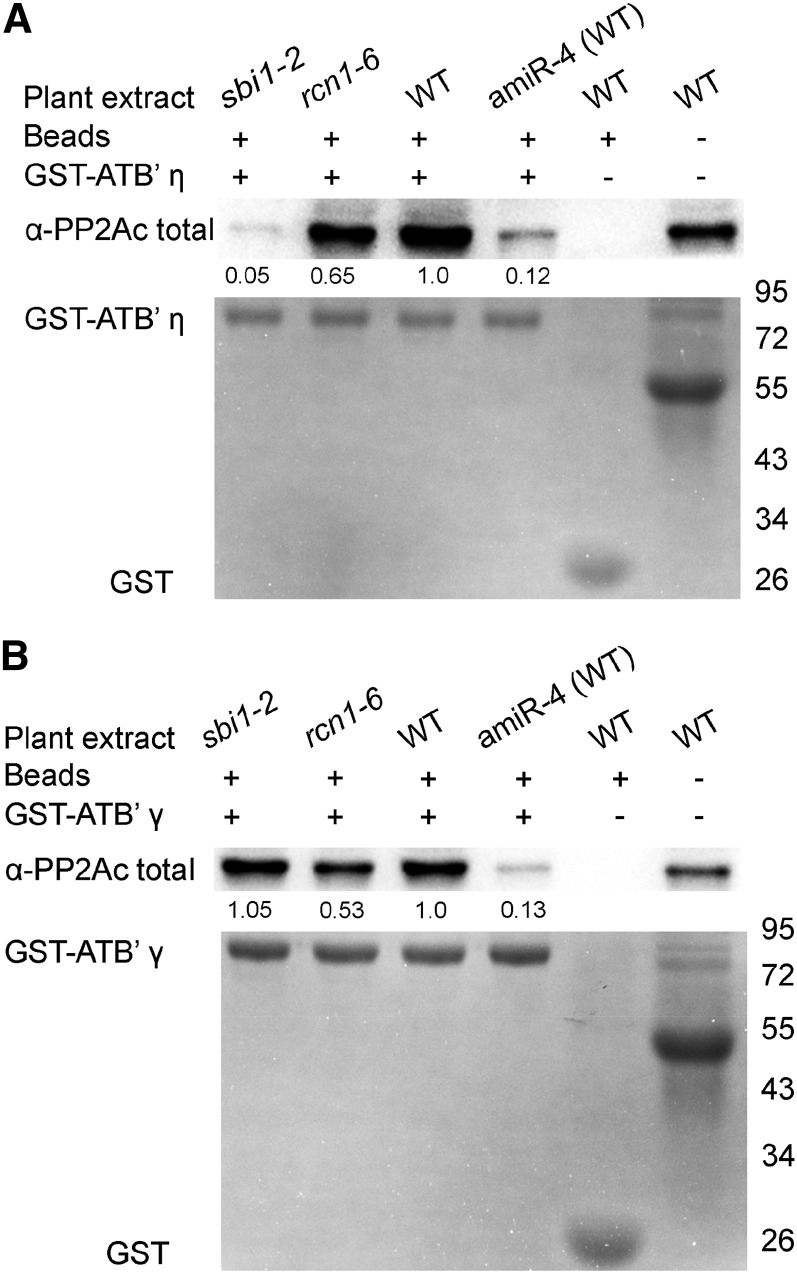
PROTEIN PHOSPHATASE 2A (PP2A) is a major group of serine/threonine protein phosphatases in eukaryotes. It is composed of three subunits: scaffolding subunit A, regulatory subunit B, and catalytic subunit C. Assembly of the PP2A holoenzyme in Arabidopsis (Arabidopsis thaliana) depends on Arabidopsis PHOSPHOTYROSYL PHOSPHATASE ACTIVATOR (AtPTPA). Reduced expression of AtPTPA leads to severe defects in plant development, altered responses to abscisic acid, ethylene, and sodium chloride, and decreased PP2A activity. In particular, AtPTPA deficiency leads to decreased methylation in PP2A-C subunits (PP2Ac). Complete loss of PP2Ac methylation in the suppressor of brassinosteroid insensitive1 mutant leads to 30% reduction of PP2A activity, suggesting that PP2A with a methylated C subunit is more active than PP2A with an unmethylated C subunit. Like AtPTPA, PP2A-A subunits are also required for PP2Ac methylation. The interaction between AtPTPA and PP2Ac is A subunit dependent. In addition, AtPTPA deficiency leads to reduced interactions of B subunits with C subunits, resulting in reduced functional PP2A holoenzyme formation. Thus, AtPTPA is a critical factor for committing the subunit A/subunit C dimer toward PP2A heterotrimer formation.
© 2014 American Society of Plant Biologists. All Rights Reserved.
Figures








Similar articles
-
Overexpression of AtPTPA in Arabidopsis increases protein phosphatase 2A activity by promoting holoenzyme formation and ABA negatively affects holoenzyme formation.Plant Signal Behav. 2015;10(11):e1052926. doi: 10.1080/15592324.2015.1052926. Plant Signal Behav. 2015. PMID: 26633567 Free PMC article.
-
The subfamily II catalytic subunits of protein phosphatase 2A (PP2A) are involved in cortical microtubule organization.Planta. 2018 Dec;248(6):1551-1567. doi: 10.1007/s00425-018-3000-0. Epub 2018 Sep 6. Planta. 2018. PMID: 30191298
-
Overexpression of PP2A-C5 that encodes the catalytic subunit 5 of protein phosphatase 2A in Arabidopsis confers better root and shoot development under salt conditions.Plant Cell Environ. 2017 Jan;40(1):150-164. doi: 10.1111/pce.12837. Epub 2016 Oct 26. Plant Cell Environ. 2017. PMID: 27676158
-
Protein Phosphatase 2A: a Double-Faced Phosphatase of Cellular System and Its Role in Neurodegenerative Disorders.Mol Neurobiol. 2018 Feb;55(2):1750-1761. doi: 10.1007/s12035-017-0444-3. Epub 2017 Feb 21. Mol Neurobiol. 2018. PMID: 28224476 Review.
-
Effects of carboxyl-terminal methylation on holoenzyme function of the PP2A subfamily.Biochem Soc Trans. 2020 Oct 30;48(5):2015-2027. doi: 10.1042/BST20200177. Biochem Soc Trans. 2020. PMID: 33125487 Free PMC article. Review.
Cited by
-
MID1-PP2A complex functions as new insights in human lung adenocarcinoma.J Cancer Res Clin Oncol. 2018 May;144(5):855-864. doi: 10.1007/s00432-018-2601-0. Epub 2018 Feb 15. J Cancer Res Clin Oncol. 2018. PMID: 29450633 Free PMC article.
-
Specific PP2A Catalytic Subunits Are a Prerequisite for Positive Growth Effects in Arabidopsis Co-Cultivated with Azospirillum brasilense and Pseudomonas simiae.Plants (Basel). 2020 Dec 30;10(1):66. doi: 10.3390/plants10010066. Plants (Basel). 2020. PMID: 33396893 Free PMC article.
-
PP2A-3 interacts with ACR4 and regulates formative cell division in the Arabidopsis root.Proc Natl Acad Sci U S A. 2016 Feb 2;113(5):1447-52. doi: 10.1073/pnas.1525122113. Epub 2016 Jan 20. Proc Natl Acad Sci U S A. 2016. PMID: 26792519 Free PMC article.
-
Mutual Regulation of Receptor-Like Kinase SIT1 and B'κ-PP2A Shapes the Early Response of Rice to Salt Stress.Plant Cell. 2019 Sep;31(9):2131-2151. doi: 10.1105/tpc.18.00706. Epub 2019 Jun 20. Plant Cell. 2019. PMID: 31221736 Free PMC article.
-
PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii.Cells. 2025 Jun 3;14(11):835. doi: 10.3390/cells14110835. Cells. 2025. PMID: 40498010 Free PMC article.
References
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR. (1999) EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
-
- Ballesteros I, Domínguez T, Sauer M, Paredes P, Duprat A, Rojo E, Sanmartín M, Sánchez-Serrano JJ. (2013) Specialized functions of the PP2A subfamily II catalytic subunits PP2A-C3 and PP2A-C4 in the distribution of auxin fluxes and development in Arabidopsis. Plant J 73: 862–872 - PubMed
-
- Bradford MM. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
