

Linking genetic variation in human Toll-like receptor 5 genes to the gut microbiome's potential to cause inflammation
- PMID: 25284610
- PMCID: PMC4259828
- DOI: 10.1016/j.imlet.2014.07.017
Linking genetic variation in human Toll-like receptor 5 genes to the gut microbiome's potential to cause inflammation
Abstract
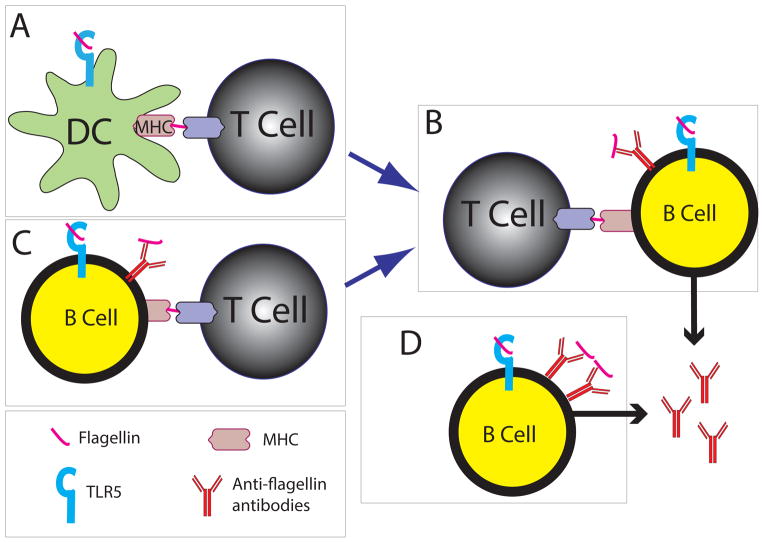
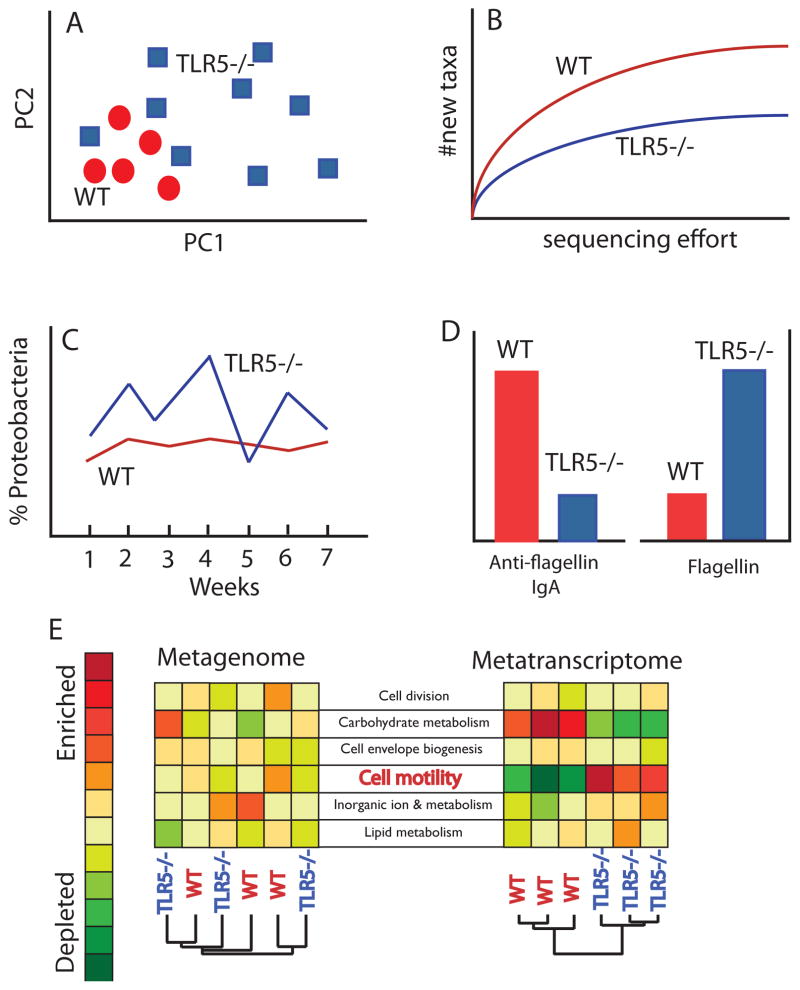
Immunodeficiencies can lead to alterations of the gut microbiome that render it pathogenic and capable of transmitting disease to naïve hosts. Here, we review the role of Toll-like receptor (TLR) 5, the innate receptor for bacterial flagellin, in immune responses to the normal gut microbiota with a focus its role on adaptive immunity. Loss of TLR5 has profound effects on the microbiota that include greater temporal instability of major lineages and upregulation of flagellar motility genes that may be linked to the reduced levels of anti-flagellin antibodies in the TLR5(-/-) host. A variety of human TLR5 gene alleles exist that also associated with inflammatory conditions and may do so via effects on the gut microbiome and altered host-microbial crosstalk.
Keywords: Adaptive immunity; Allelic variation; Gut microbiome; Metagenomics; TLR5; Toll-like receptor 5.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
