

On the Origin of the Canonical Nucleobases: An Assessment of Selection Pressures across Chemical and Early Biological Evolution
- PMID: 25284884
- PMCID: PMC4181368
- DOI: 10.1002/ijch.201300009
On the Origin of the Canonical Nucleobases: An Assessment of Selection Pressures across Chemical and Early Biological Evolution
Abstract
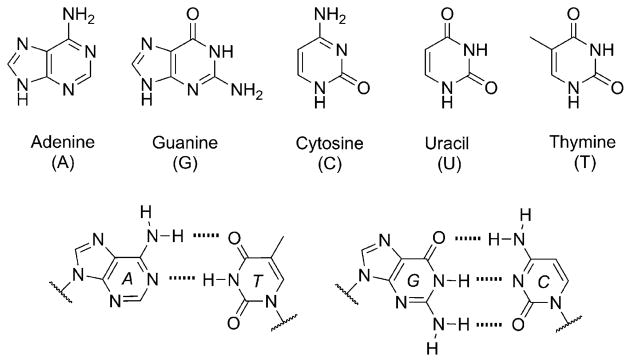
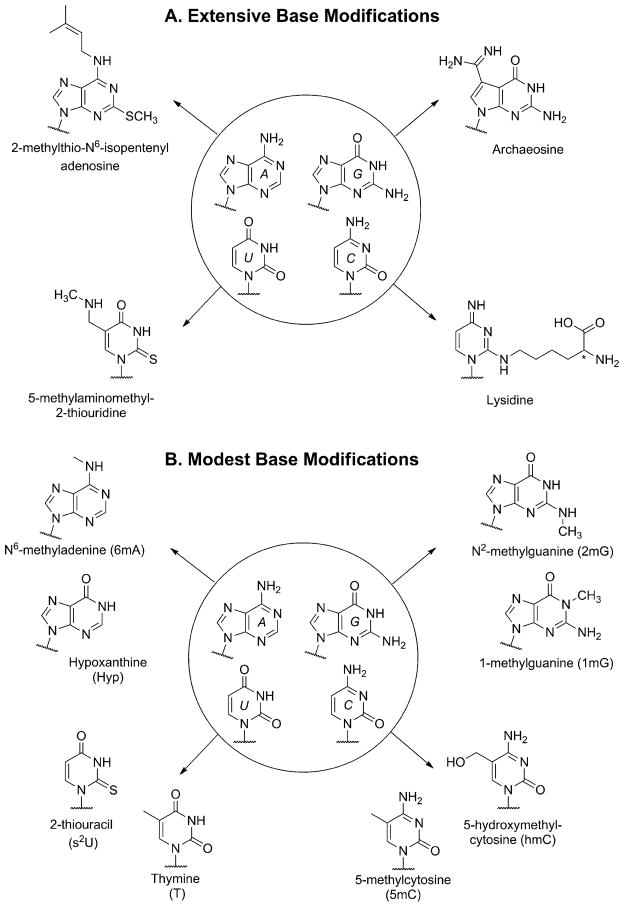
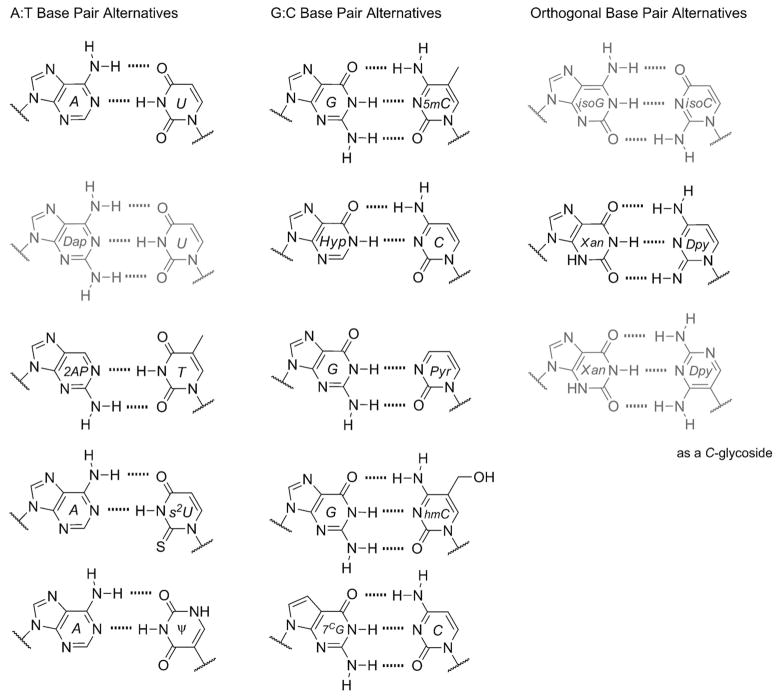
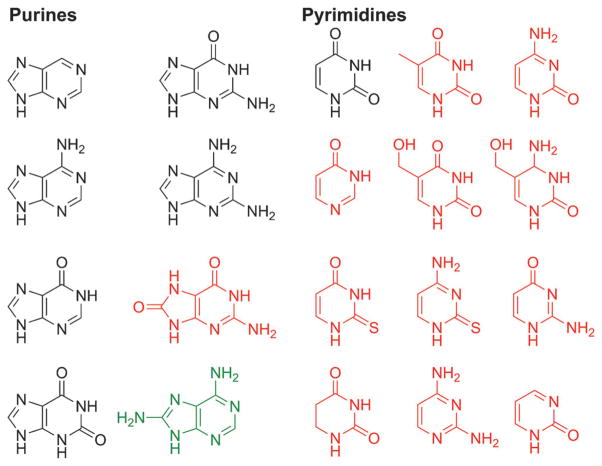
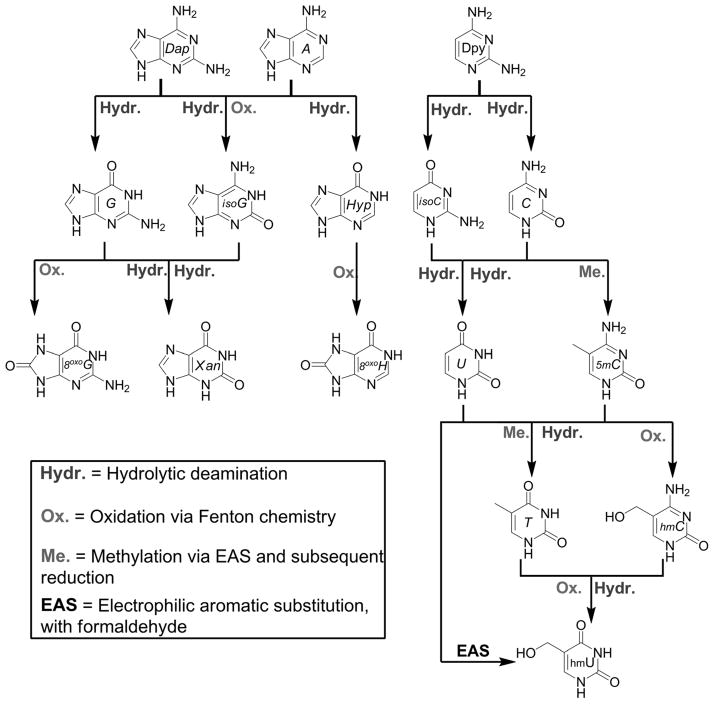
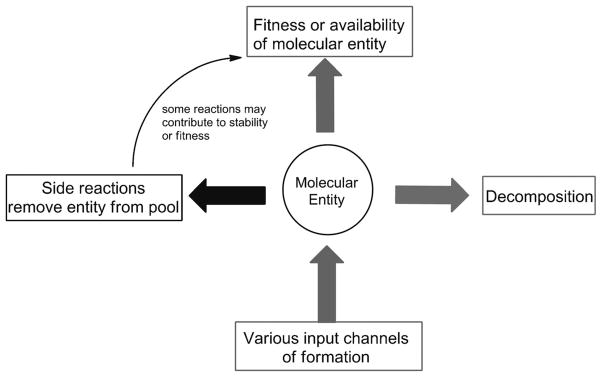
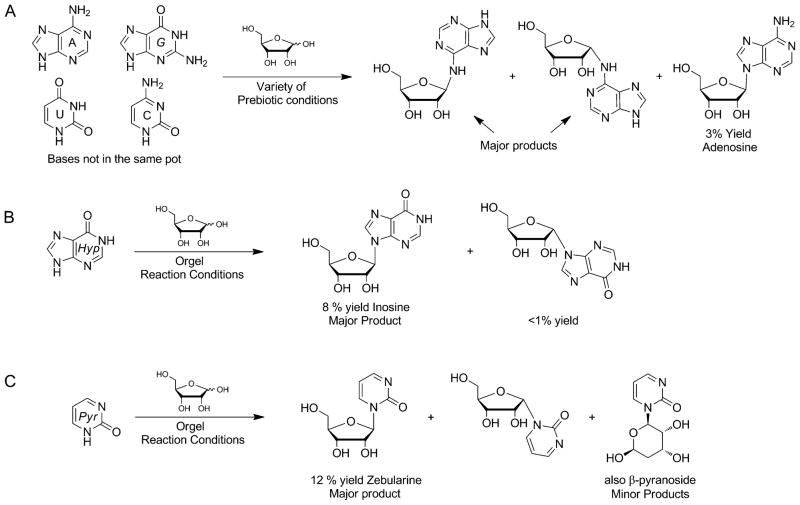
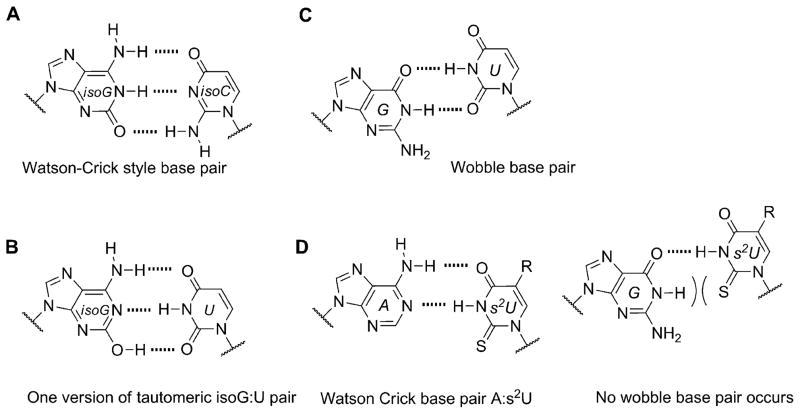
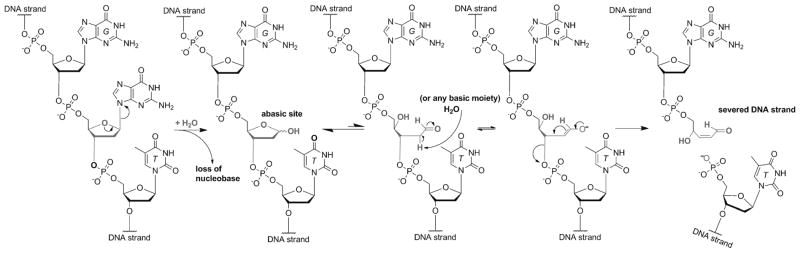
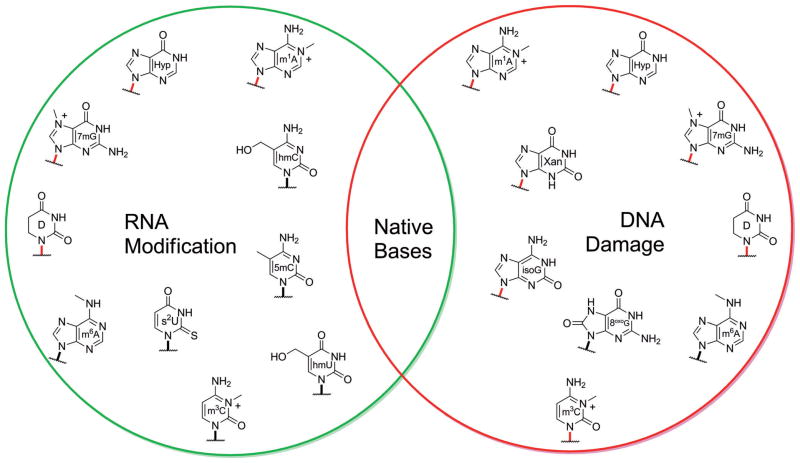
The native bases of RNA and DNA are prominent examples of the narrow selection of organic molecules upon which life is based. How did nature "decide" upon these specific heterocycles? Evidence suggests that many types of heterocycles could have been present on the early Earth. It is therefore likely that the contemporary composition of nucleobases is a result of multiple selection pressures that operated during early chemical and biological evolution. The persistence of the fittest heterocycles in the prebiotic environment towards, for example, hydrolytic and photochemical assaults, may have given some nucleobases a selective advantage for incorporation into the first informational polymers. The prebiotic formation of polymeric nucleic acids employing the native bases remains, however, a challenging problem to reconcile. Hypotheses have proposed that the emerging RNA world may have included many types of nucleobases. This is supported by the extensive utilization of non-canonical nucleobases in extant RNA and the resemblance of many of the modified bases to heterocycles generated in simulated prebiotic chemistry experiments. Selection pressures in the RNA world could have therefore narrowed the composition of the nucleic acid bases. Two such selection pressures may have been related to genetic fidelity and duplex stability. Considering these possible selection criteria, the native bases along with other related heterocycles seem to exhibit a certain level of fitness. We end by discussing the strength of the N-glycosidic bond as a potential fitness parameter in the early DNA world, which may have played a part in the refinement of the alphabetic bases.
Keywords: DNA; RNA world; molecular evolution; nucleobases; prebiotic chemistry.
Figures

References
-
- Carell T, Brandmayr C, Hienzsch A, Müller M, Pearson D, Reiter V, Thoma I, Thumbs P, Wagner M. Angew Chem, Int Ed. 2012;51:7110. - PubMed
-
- Martínez Giménez JA, Sáez GT, Seisdedos RT. J Theor Biol. 1998;194:485. - PubMed
- Forterre P, Grosjean H. In: DNA and RNA Modification Enzymes: Structure, Mechanism, Function and Evolution. Grosjean H, editor. Landes Bioscience; Austin, Texas: 2009. p. 259.
- Cermakian N, Cedergren R. In: Modification and Editing of RNA. Grosjean H, Benne R, editors. ASM Press; Washington, D. C: 1998. p. 535.
-
- Berg JM, Tymoczko JL, Stryer L. Biochemistry. 6. W. H. Freeman; New York: 2007.
-
- Poole A, Penny D, Sjöberg BM. Nat Rev Mol Cell Biol. 2001;2:147. - PubMed
-
- Warren RAJ. Annu Rev Microbiol. 1980;34:137. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources