

Insights into the binding of PARP inhibitors to the catalytic domain of human tankyrase-2
- PMID: 25286857
- PMCID: PMC4188013
- DOI: 10.1107/S1399004714017660
Insights into the binding of PARP inhibitors to the catalytic domain of human tankyrase-2
Abstract
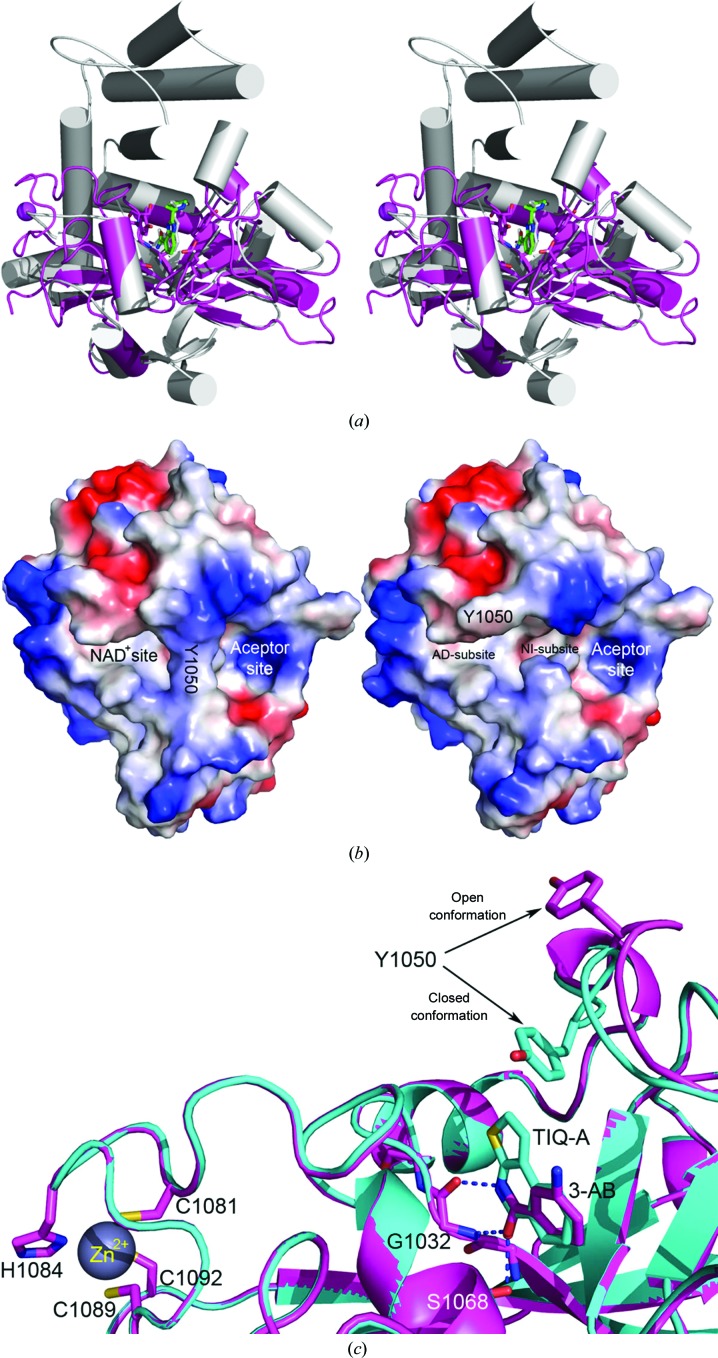
The poly(ADP-ribose) polymerase (PARP) family represents a new class of therapeutic targets with diverse potential disease indications. PARP1 and PARP2 inhibitors have been developed for breast and ovarian tumors manifesting double-stranded DNA-repair defects, whereas tankyrase 1 and 2 (TNKS1 and TNKS2, also known as PARP5a and PARP5b, respectively) inhibitors have been developed for tumors with elevated β-catenin activity. As the clinical relevance of PARP inhibitors continues to be actively explored, there is heightened interest in the design of selective inhibitors based on the detailed structural features of how small-molecule inhibitors bind to each of the PARP family members. Here, the high-resolution crystal structures of the human TNKS2 PARP domain in complex with 16 various PARP inhibitors are reported, including the compounds BSI-201, AZD-2281 and ABT-888, which are currently in Phase 2 or 3 clinical trials. These structures provide insight into the inhibitor-binding modes for the tankyrase PARP domain and valuable information to guide the rational design of future tankyrase-specific inhibitors.
Keywords: TNKS2; cancer; poly(ADP-ribose) polymerase; structural biology; structure-based drug discovery.
Figures





Similar articles
-
Structural Basis for Potency and Promiscuity in Poly(ADP-ribose) Polymerase (PARP) and Tankyrase Inhibitors.J Med Chem. 2017 Feb 23;60(4):1262-1271. doi: 10.1021/acs.jmedchem.6b00990. Epub 2016 Dec 21. J Med Chem. 2017. PMID: 28001384 Free PMC article.
-
Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors.Nat Biotechnol. 2012 Feb 19;30(3):283-8. doi: 10.1038/nbt.2121. Nat Biotechnol. 2012. PMID: 22343925
-
Structural basis of selective inhibition of human tankyrases.J Med Chem. 2012 Feb 9;55(3):1360-7. doi: 10.1021/jm201510p. Epub 2012 Jan 25. J Med Chem. 2012. PMID: 22233320
-
The antimitotic potential of PARP inhibitors, an unexplored therapeutic alternative.Curr Top Med Chem. 2014;14(20):2346-65. doi: 10.2174/1568026614666141130100641. Curr Top Med Chem. 2014. PMID: 25434352 Review.
-
Insights of tankyrases: A novel target for drug discovery.Eur J Med Chem. 2020 Dec 1;207:112712. doi: 10.1016/j.ejmech.2020.112712. Epub 2020 Aug 17. Eur J Med Chem. 2020. PMID: 32877803 Review.
Cited by
-
MicroRNA-206 inhibits influenza A virus replication by targeting tankyrase 2.Cell Microbiol. 2021 Feb;23(2):e13281. doi: 10.1111/cmi.13281. Epub 2020 Nov 4. Cell Microbiol. 2021. PMID: 33099847 Free PMC article.
-
Molecular Mechanism of Selective Binding of NMS-P118 to PARP-1 and PARP-2: A Computational Perspective.Front Mol Biosci. 2020 Apr 15;7:50. doi: 10.3389/fmolb.2020.00050. eCollection 2020. Front Mol Biosci. 2020. PMID: 32373627 Free PMC article.
-
PARP inhibition in leukocytes diminishes inflammation via effects on integrins/cytoskeleton and protects the blood-brain barrier.J Neuroinflammation. 2016 Sep 27;13(1):254. doi: 10.1186/s12974-016-0729-x. J Neuroinflammation. 2016. PMID: 27677851 Free PMC article.
-
Targeting PARP proteins in acute leukemia: DNA damage response inhibition and therapeutic strategies.J Hematol Oncol. 2022 Jan 22;15(1):10. doi: 10.1186/s13045-022-01228-0. J Hematol Oncol. 2022. PMID: 35065680 Free PMC article. Review.
-
Structural basis of tankyrase activation by polymerization.Nature. 2022 Dec;612(7938):162-169. doi: 10.1038/s41586-022-05449-8. Epub 2022 Nov 23. Nature. 2022. PMID: 36418402 Free PMC article.
References
-
- Adams, P. D. et al. (2010). Acta Cryst. D66, 213–221. - PubMed
-
- Ahel, I., Ahel, D., Matsusaka, T., Clark, A. J., Pines, J., Boulton, S. J. & West, S. C. (2008). Nature (London), 451, 81–85. - PubMed
-
- Bauer, P. I., Kirsten, E., Varadi, G., Young, L. J., Hakam, A., Comstock, J. A. & Kun, E. (1995). Biochimie, 77, 374–377. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
