

Comparative evaluation of lesion development, tissue damage, and cytokine expression in golden hamsters (Mesocricetus auratus) infected by inocula with different Leishmania (Viannia) braziliensis concentrations
- PMID: 25287925
- PMCID: PMC4249292
- DOI: 10.1128/IAI.02083-14
Comparative evaluation of lesion development, tissue damage, and cytokine expression in golden hamsters (Mesocricetus auratus) infected by inocula with different Leishmania (Viannia) braziliensis concentrations
Abstract
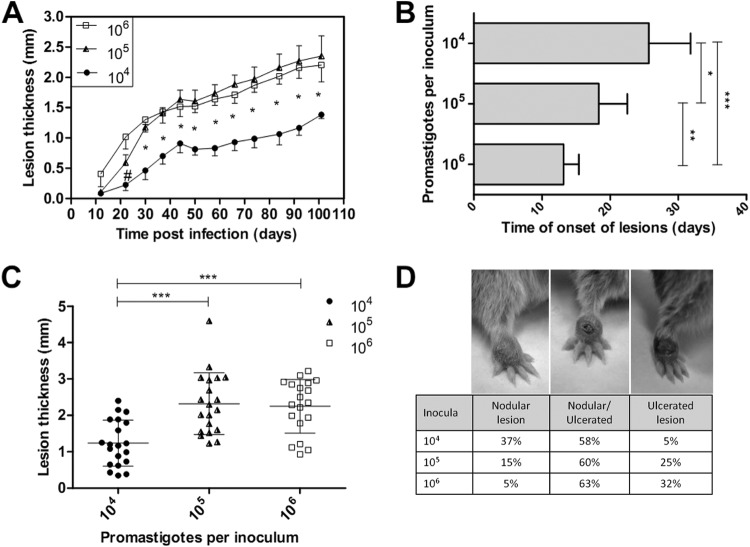
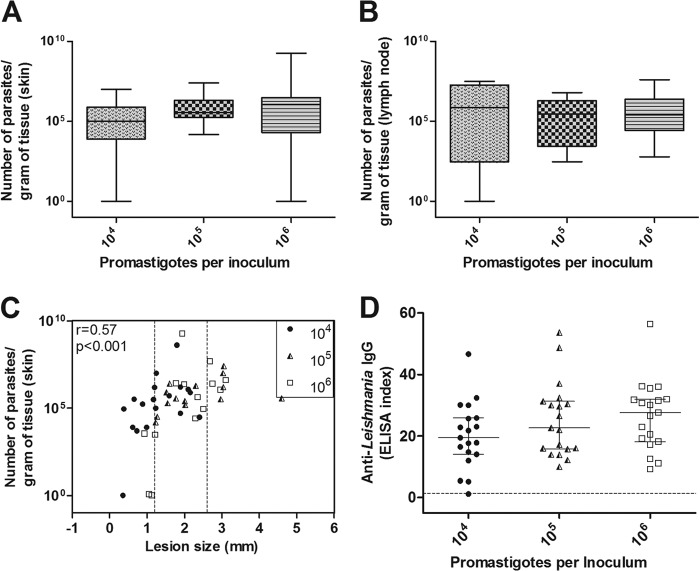
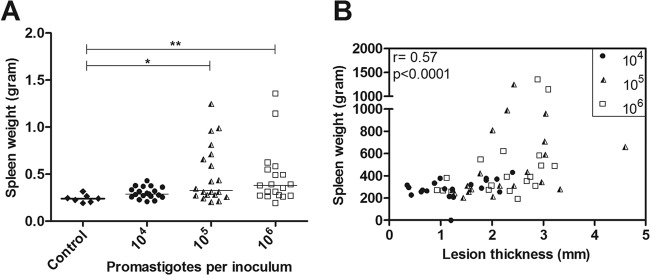
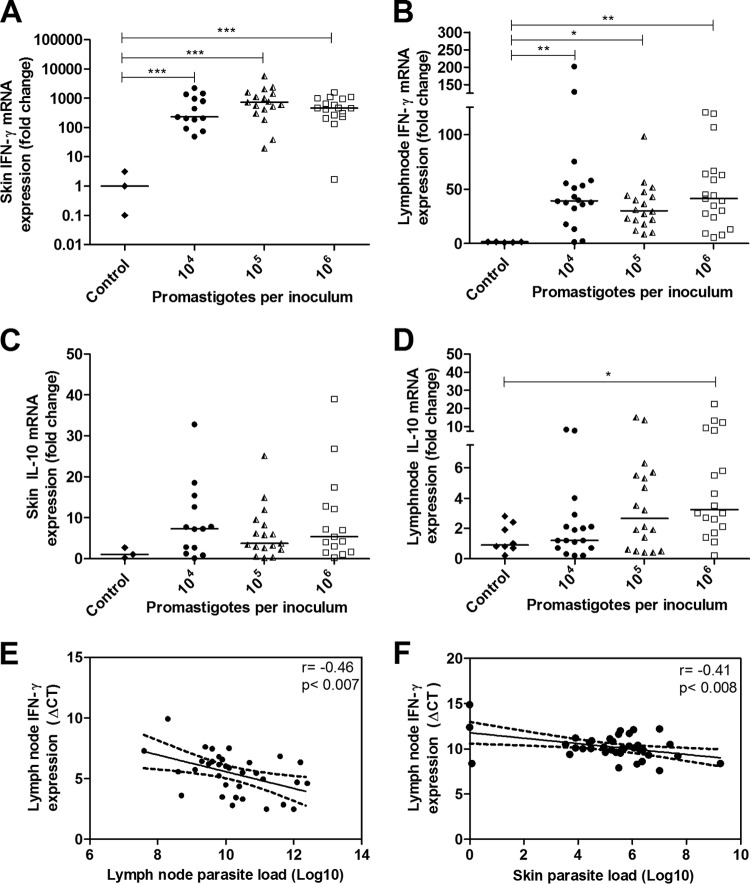
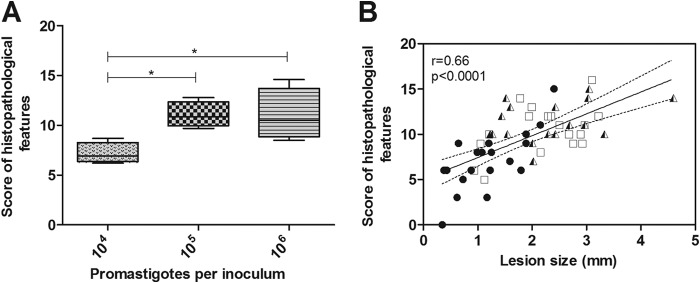
The golden hamster (Mesocricetus auratus) is a susceptible model to Leishmania (Viannia) spp.; however, available studies employ different infection protocols, which account for clinical and pathological presentation differences. Herein, L. (V.) braziliensis preparations were standardized to contain 10(4), 10(5), or 10(6) parasites to determine an optimal inoculum that ensured cutaneous lesions without causing a disseminated infection in hamsters. Lesion development was followed for 105 days by size measurements, and skin, draining lymph node, spleen, and sera were investigated to check parasite load, spleen visceralization, cytokine expression, histopathological changes, and anti-Leishmania IgG levels. The lesion emergence time was inversely proportional to the parasite concentration in the inocula. Animals infected by 10(4) parasites presented nodular lesions, while those infected with 10(6) parasites often exhibited ulcerated lesions. The differences in the final lesion sizes were observed between 10(4) and 10(5) inocula or 10(4) and 10(6) inocula. High IFNG expression, anti-Leishmania IgG levels, and parasite load occurred independently of the inoculum used. A mild inflammatory skin involvement was observed in animals infected with 10(4) parasites, while extensive tissue damage and parasite spleen visceralization occurred with 10(5) and 10(6) parasites. These results indicate that inocula with different concentrations of parasites generate differences in the time of lesion emergence, clinical presentation, and systemic commitment, despite high and similar IFNG expression and parasite load. This suggests that a modulation in the immune response to different parasite numbers occurs in an early phase of the infection, which could dictate the establishment and magnitude of the chronic phase of the disease.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures

References
-
- Schriefer A, Schriefer AL, Góes-Neto A, Gumarães LH, Carvalho LP, Almeida RP, Machado PR, Lessa HÁ de Jesus AR, Riley LW, Carvalho EM. 2004. Multiclonal Leishmania braziliensis population structure and its clinical implication in a region of endemicity for American cutaneous leishmaniasis. Infect. Immun. 72:508–514. 10.1128/IAI.72.1.508-514.2004. - DOI - PMC - PubMed
-
- Costa JM, Saldanha AC, Nascimento D, Sampaio G, Carneiro F, Lisboa E, Silva ML, Barral A. 2009. Clinical modalities, diagnosis and therapeutic approach of the tegumentary leishmaniasis in Brazil. GM Bahia 79:70–83.
-
- Cupolillo E, Brahim LR, Toaldo CB, de Oliveira-Neto MP, de Brito ME, Falqueto A, de Farias Naiff M, Grimaldi G., Jr 2003. Genetic polymorphism and molecular epidemiology of Leishmania (Viannia) braziliensis from different hosts and geographic areas in Brasil. J. Clin. Microbiol. 41:3126–3132. 10.1128/JCM.41.7.3126-3132.2003. - DOI - PMC - PubMed
-
- Coutinho S, Pirmez C, Mendonça S, Conceição-Silva F, Dórea R. 1987. Pathogenesis in immunopathology of leishmaniasis. Mem. Inst. Oswaldo Cruz 82:214–228. 10.1590/S0074-02761987000500005. - DOI
-
- Jeronimo SM, Duggal P, Ettinger NA, Nascimento ET, Monteiro GR, Cabral AP, Pontes NN, Lacerda HG, Queiroz PV, Gomes CE, Pearson RD, Blackwell JM, Beaty TH, Wilson ME. 2007. Genetic predisposition to self-curing infection with the protozoan Leishmania chagasi: a genomewide scan. J. Infect. Dis. 196:1261–1269. 10.1086/521682. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
