

Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein
- PMID: 25288733
- PMCID: PMC4210292
- DOI: 10.1073/pnas.1407087111
Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein
Abstract
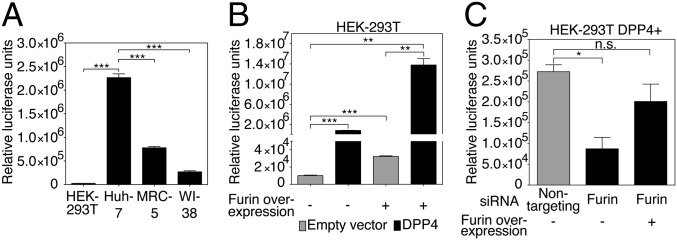
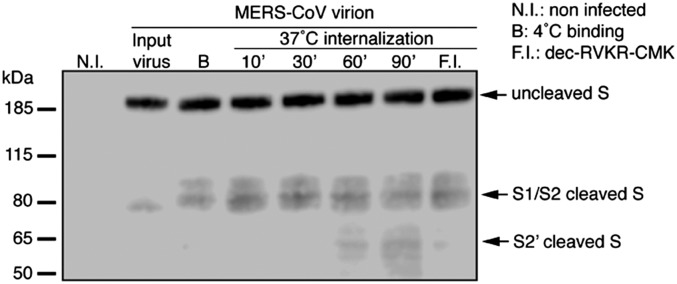
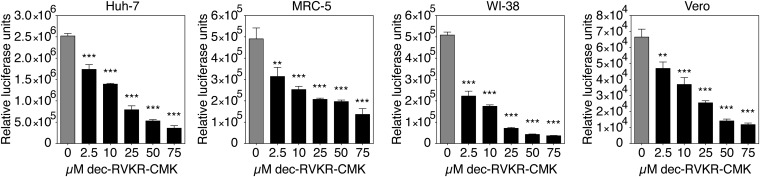
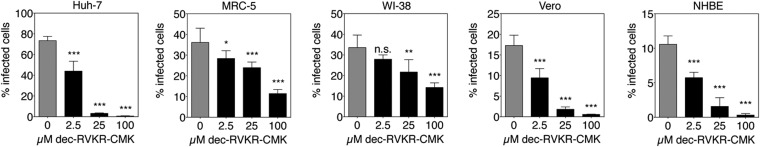
Middle East respiratory syndrome coronavirus (MERS-CoV) is a newly identified betacoronavirus causing high morbidity and mortality in humans. The coronavirus spike (S) protein is the main determinant of viral entry, and although it was previously shown that MERS-CoV S can be activated by various proteases, the details of the mechanisms of proteolytic activation of fusion are still incompletely characterized. Here, we have uncovered distinctive characteristics of MERS-CoV S. We identify, by bioinformatics and peptide cleavage assays, two cleavage sites for furin, a ubiquitously expressed protease, which are located at the S1/S2 interface and at the S2' position of the S protein. We show that although the S1/S2 site is proteolytically processed by furin during protein biosynthesis, the S2' site is cleaved upon viral entry. MERS-CoV pseudovirion infection was shown to be enhanced by elevated levels of furin expression, and entry could be decreased by furin siRNA silencing. Enhanced furin activity appeared to partially override the low pH-dependent nature of MERS-CoV entry. Inhibition of furin activity was shown to decrease MERS-CoV S-mediated entry, as well as infection by the virus. Overall, we show that MERS-CoV has evolved an unusual two-step furin activation for fusion, suggestive of a role during the process of emergence into the human population. The ability of MERS-CoV to use furin in this manner, along with other proteases, may explain the polytropic nature of the virus.
Keywords: Middle East respiratory syndrome coronavirus; furin; proteolytic activation; spike protein; virus entry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Zaki AM, van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367(19):1814–1820. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
