

Transcriptional coactivator CBP upregulates hTERT expression and tumor growth and predicts poor prognosis in human lung cancers
- PMID: 25294805
- PMCID: PMC4253439
- DOI: 10.18632/oncotarget.2430
Transcriptional coactivator CBP upregulates hTERT expression and tumor growth and predicts poor prognosis in human lung cancers
Abstract
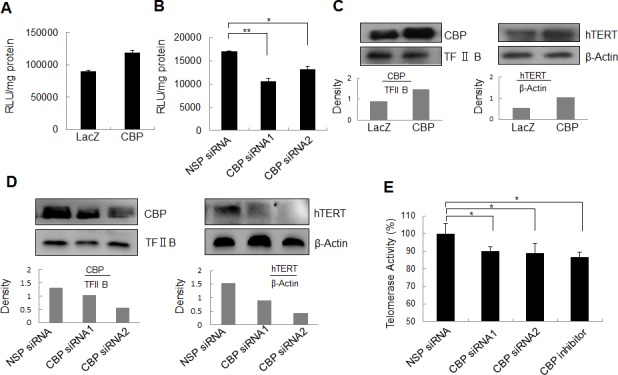
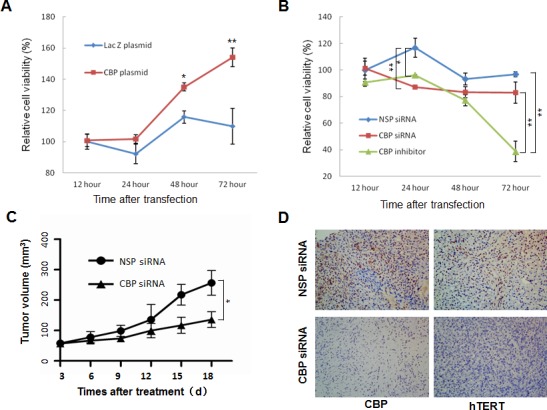
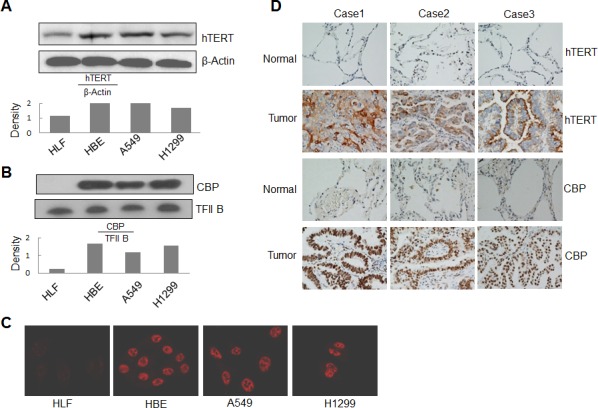
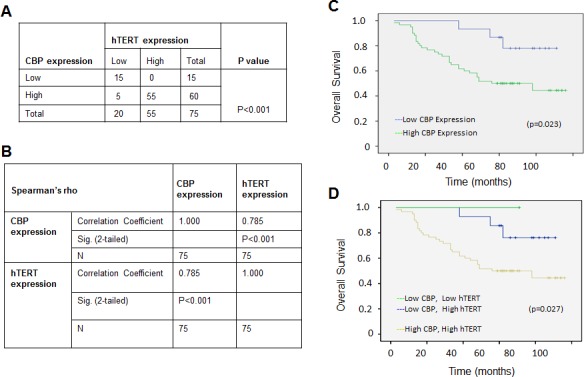
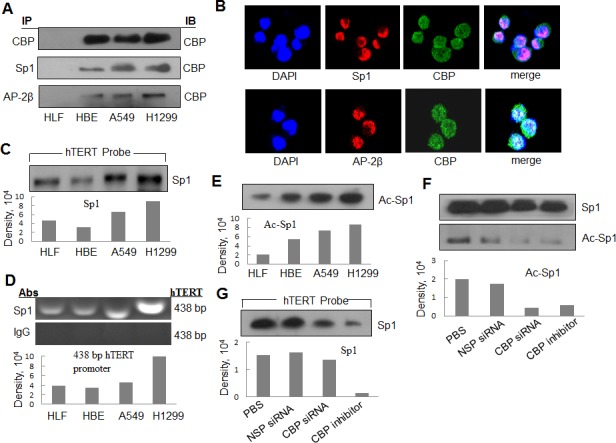
Upregulated expression and activation of human telomerase reverse transcriptase (hTERT) is a hallmarker of lung tumorigenesis. However, the mechanism underlying the aberrant hTERT activity in lung cancer cells remains poorly understood. In this study, we found the transcriptional co-activator CBP as a new hTERT promoter-binding protein that regulated hTERT expression and tumor growth in lung adenocarcinoma cells using a biotin-streptavidin-bead pulldown technique. Chromatin immunoprecipitation assay verified the immortalized cell and tumor cell-specific binding of CBP on hTERT promoter. Overexpression of exogenous CBP upregulated the expression of the hTERT promoter-driven luciferase and endogenous hTERT protein in lung cancer cells. Conversely, inhibition of CBP by CBP-specific siRNA or its chemical inhibitor repressed the expression of hTERT promoter-driven luciferase and endogenous hTERT protein as well as telomerase activity. Moreover, inhibition of CBP expression or activity also significantly reduced the proliferation of lung cancer cells in vitro and tumor growth in an xenograft mouse model in vivo. Immunohistochemical analysis of tissue microarrays of lung cancers revealed a positive correlation between CBP and hTERT. Importantly, the patients with high CBP and hTERT expression had a significantly shorter overall survival. Furthermore, CBP was found to interact with and acetylate transactivator Sp1 in lung cancer cells. Inhibition of CBP by CBP-specific siRNA or its chemical inhibitor significantly inhibited Sp1 acetylation and its binding to the hTERT promoter. Collectively, our results indicate that CBP contributes to the upregulation of hTERT expression and tumor growth, and overexpression of CBP predicts poor prognosis in human lung cancers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Tomas-Loba A, Flores I, Fernandez-Marcos PJ, Cayuela ML, Maraver A, Tejera A, Borras C, Matheu A, Klatt P, Flores JM, Vina J, Serrano M, Blasco MA. Telomerase reverse transcriptase delays aging in cancer-resistant mice. Cell. 2008;135(4):609–622. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
