

Antinematode activity of Violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans
- PMID: 25295516
- PMCID: PMC4189955
- DOI: 10.1371/journal.pone.0109201
Antinematode activity of Violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans
Erratum in
-
Correction: Antinematode Activity of Violacein and the Role of the Insulin/IGF-1 Pathway in Controlling Violacein Sensitivity in Caenorhabditis elegans.PLoS One. 2018 Dec 26;13(12):e0210026. doi: 10.1371/journal.pone.0210026. eCollection 2018. PLoS One. 2018. PMID: 30586436 Free PMC article.
Abstract
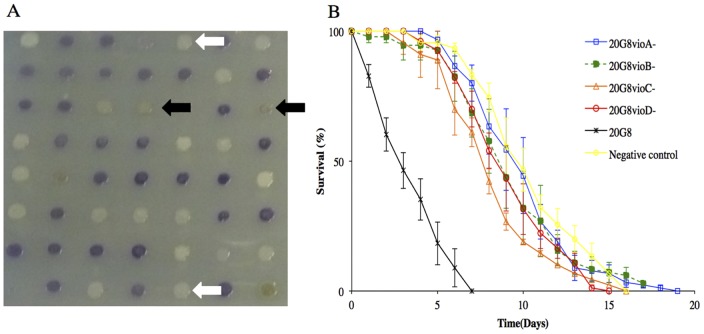
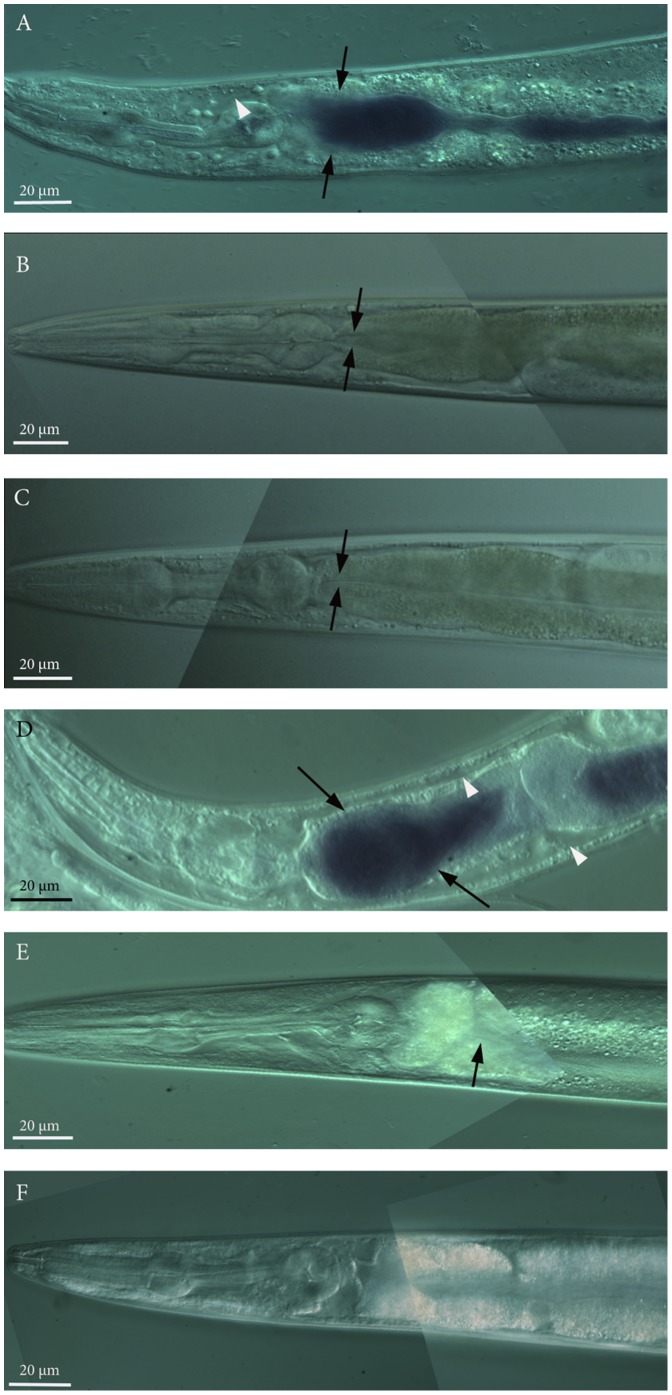
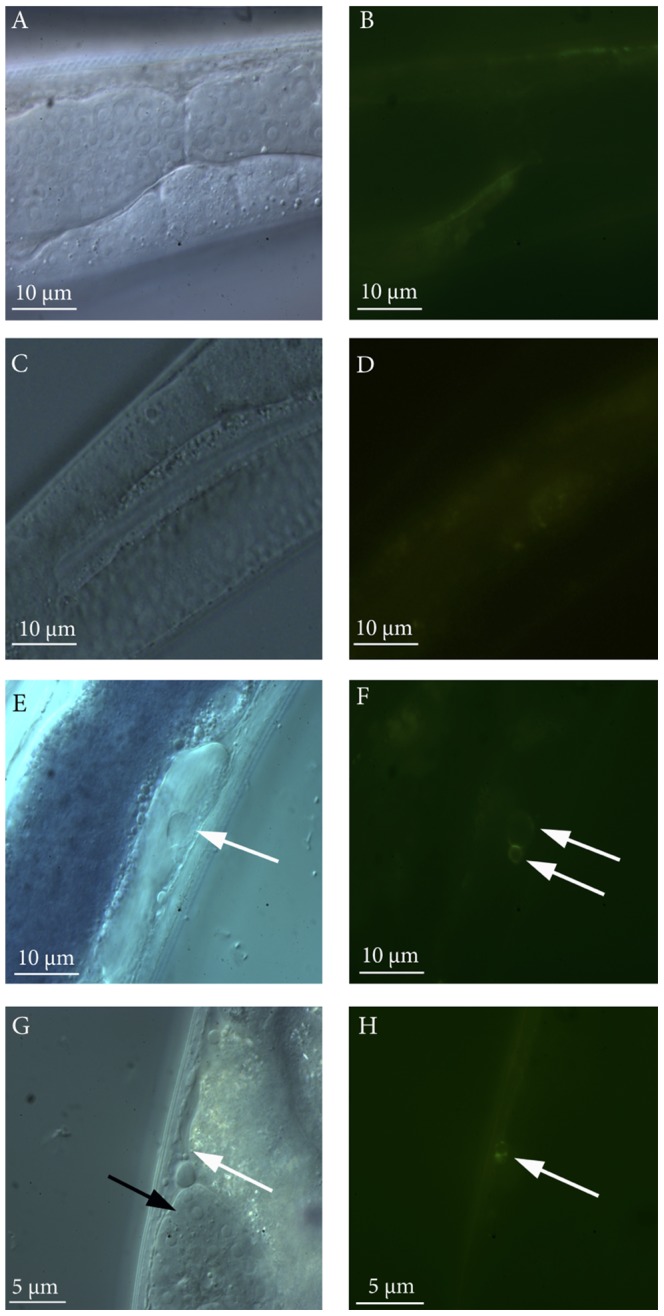
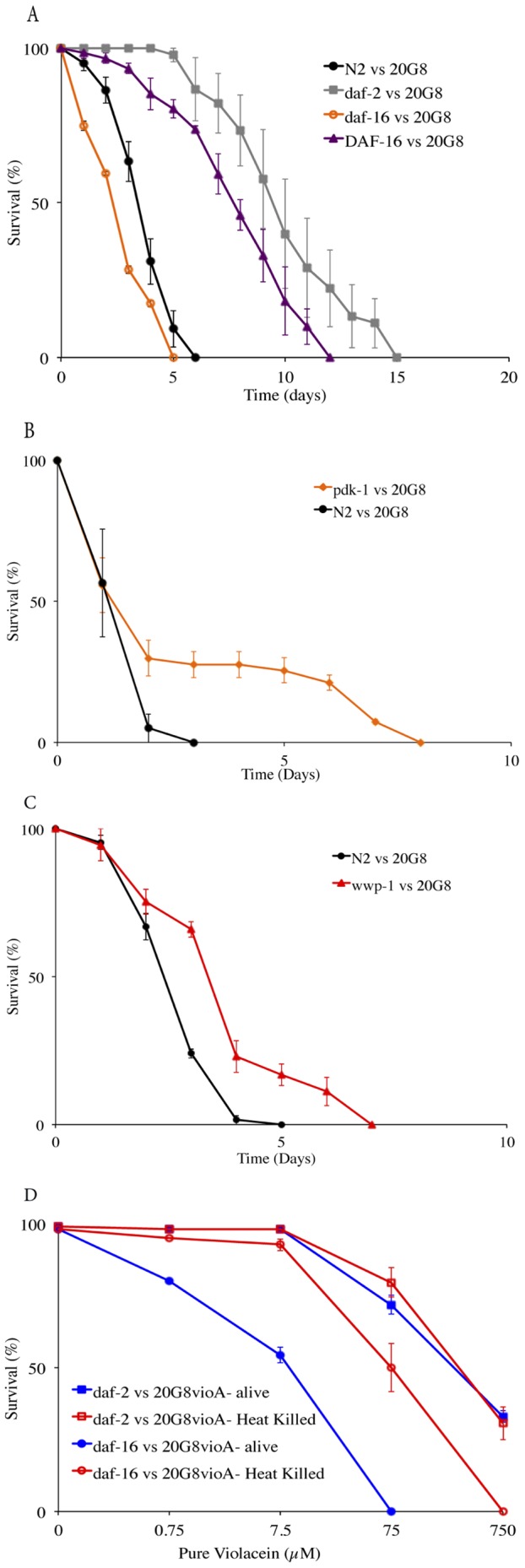
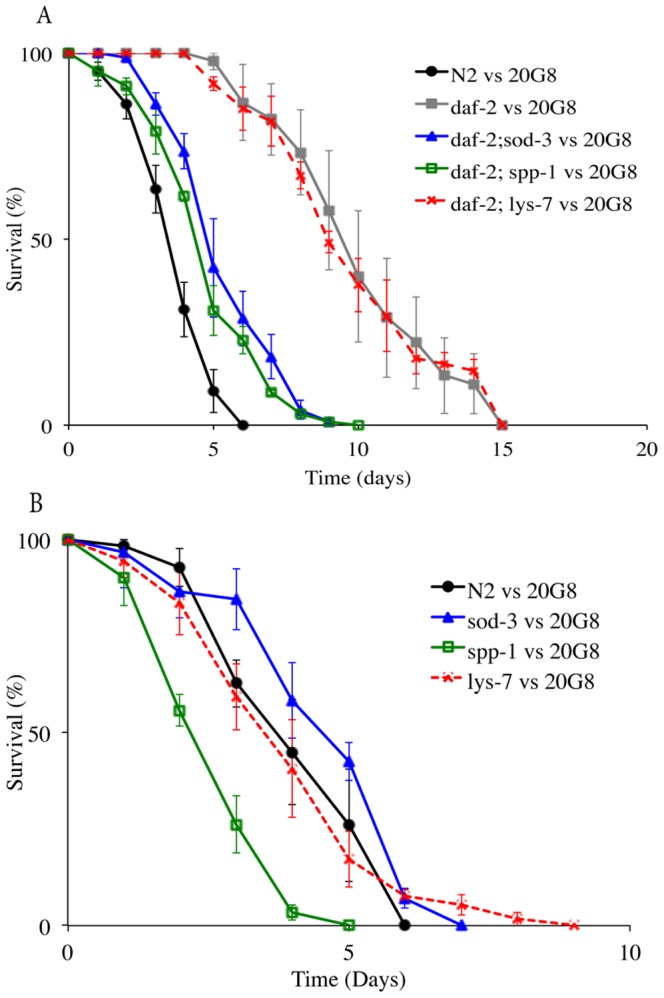
The purple pigment violacein is well known for its numerous biological activities including antibacterial, antiviral, antiprotozoan, and antitumor effects. In the current study we identify violacein as the antinematode agent produced by the marine bacterium Microbulbifer sp. D250, thereby extending the target range of this small molecule. Heterologous expression of the violacein biosynthetic pathway in E. coli and experiments using pure violacein demonstrated that this secondary metabolite facilitates bacterial accumulation in the nematode intestine, which is accompanied by tissue damage and apoptosis. Nematodes such as Caenorhabditis elegans utilise a well-defined innate immune system to defend against pathogens. Using C. elegans as a model we demonstrate the DAF-2/DAF-16 insulin/IGF-1 signalling (IIS) component of the innate immune pathway modulates sensitivity to violacein-mediated killing. Further analysis shows that resistance to violacein can occur due to a loss of DAF-2 function and/or an increased function of DAF-16 controlled genes involved in antimicrobial production (spp-1) and detoxification (sod-3). These data suggest that violacein is a novel candidate antinematode agent and that the IIS pathway is also involved in the defence against metabolites from non-pathogenic bacteria.
Conflict of interest statement
Figures

References
-
- Kaplan RM (2004) Drug resistance in nematodes of veterinary importance: a status report. Trends Parasitol 20: 477–481. - PubMed
-
- Prichard RK, Geary TG (2008) Drug discovery: Fresh hope to can the worms. Nature 452: 157–158. - PubMed
-
- Jones AK, Buckingham SD, Sattelle DB (2005) Chemistry-to-gene screens in Caenorhabditis elegans . Nat Rev Drug Discov 4: 321–330. - PubMed
-
- Sifri CD, Begun J, Ausubel FM (2005) The worm has turned-microbial virulence modeled in Caenorhabditis elegans . Trends Microbiol 13: 119–127. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
