

Determining intrachain diffusion coefficients for biopolymer dynamics from single-molecule force spectroscopy measurements
- PMID: 25296317
- PMCID: PMC4190644
- DOI: 10.1016/j.bpj.2014.08.007
Determining intrachain diffusion coefficients for biopolymer dynamics from single-molecule force spectroscopy measurements
Abstract
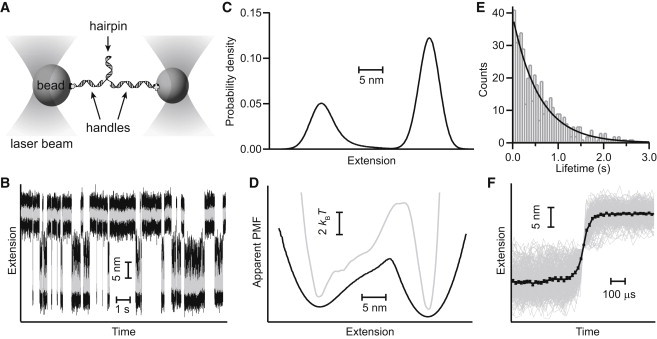
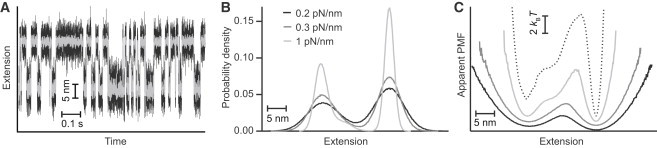
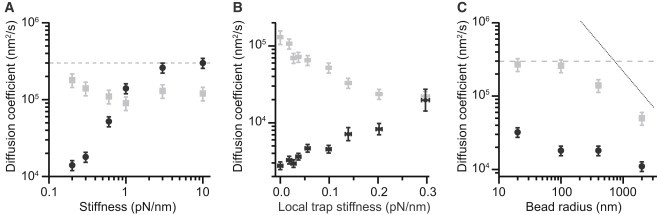
The conformational diffusion coefficient for intrachain motions in biopolymers, D, sets the timescale for structural dynamics. Recently, force spectroscopy has been applied to determine D both for unfolded proteins and for the folding transitions in proteins and nucleic acids. However, interpretation of the results remains unsettled. We investigated how instrumental effects arising from the force probes used in the measurement can affect the value of D recovered via force spectroscopy. We compared estimates of D for the folding of DNA hairpins found from measurements of rates and energy landscapes made using optical tweezers with estimates obtained from the same single-molecule trajectories via the transition path time. The apparent D obtained from the rates was much lower than the result found from the same data using transition time analysis, reflecting the effects of the mechanical properties of the force probe. Deconvolution of the finite compliance effects on the measurement allowed the intrinsic value to be recovered. These results were supported by Brownian dynamics simulations of the effects of force-probe compliance and bead size.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Dill K.A., MacCallum J.L. The protein-folding problem, 50 years on. Science. 2012;338:1042–1046. - PubMed
-
- Kramers H.A. Brownian motion in a field of force and the diffusion model of chemical reactions. Physica. 1940;7:284–304.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
