

Sensors at centrosomes reveal determinants of local separase activity
- PMID: 25299182
- PMCID: PMC4191886
- DOI: 10.1371/journal.pgen.1004672
Sensors at centrosomes reveal determinants of local separase activity
Abstract
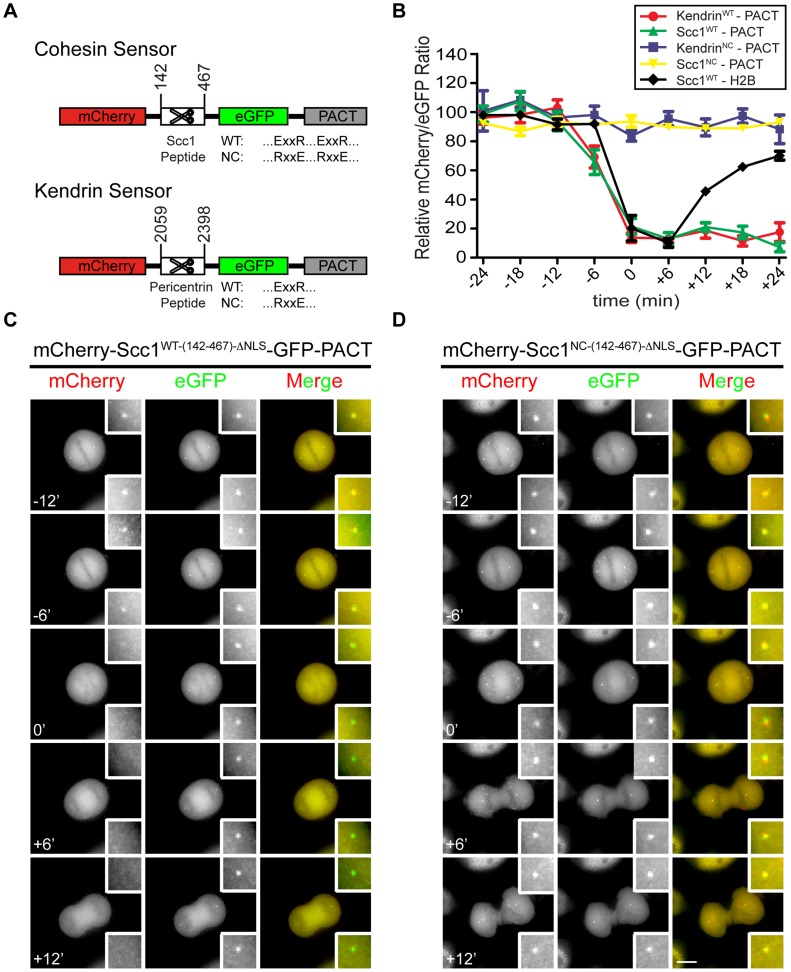
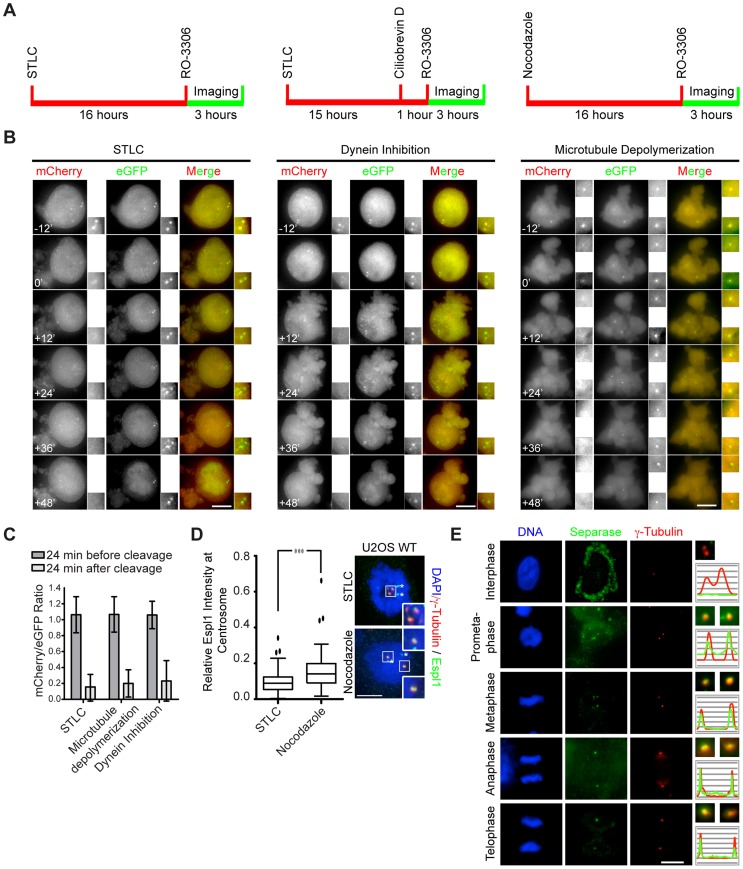
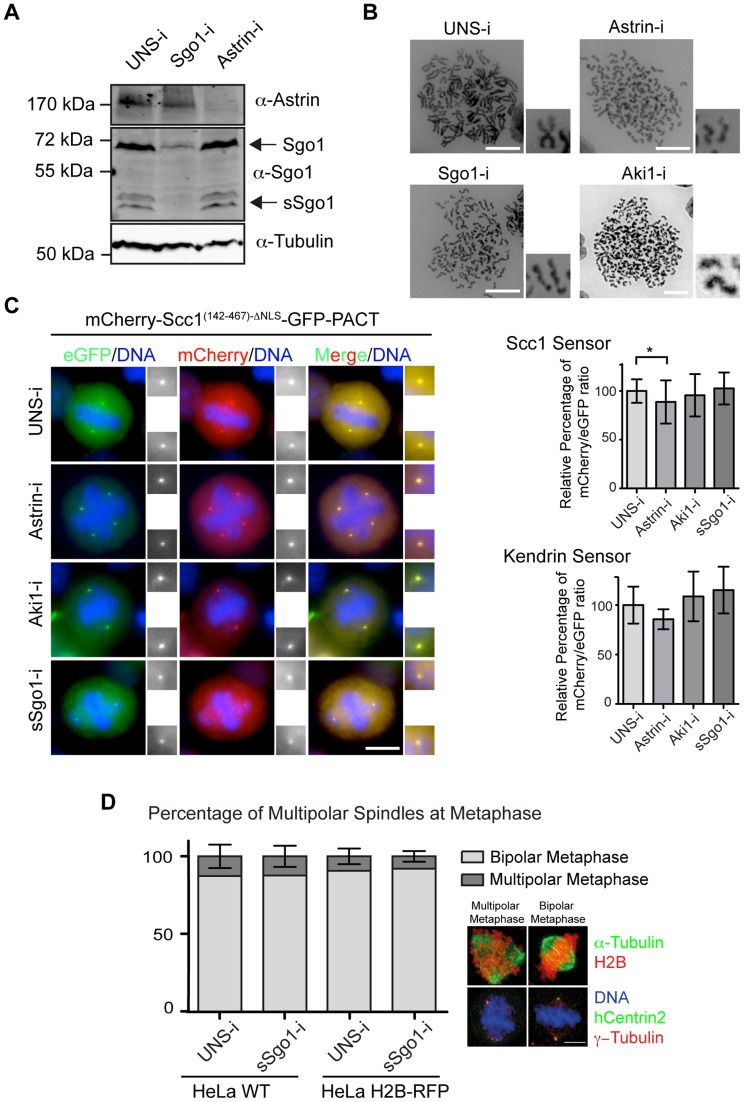
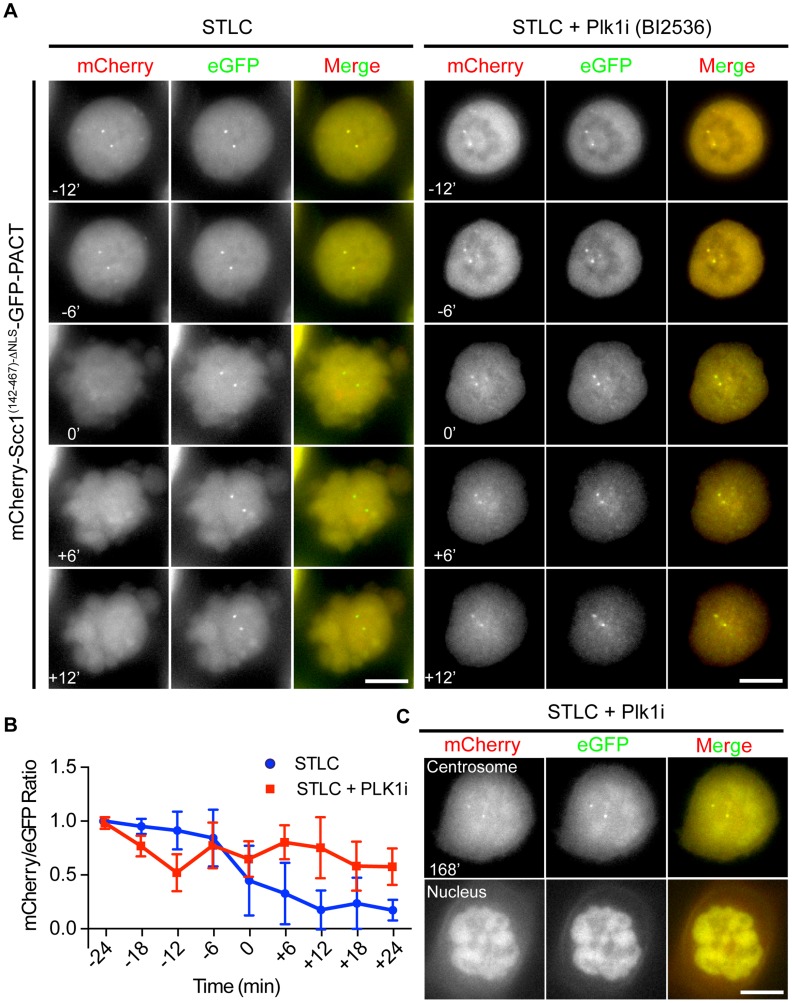
Separase is best known for its function in sister chromatid separation at the metaphase-anaphase transition. It also has a role in centriole disengagement in late mitosis/G1. To gain insight into the activity of separase at centrosomes, we developed two separase activity sensors: mCherry-Scc1(142-467)-ΔNLS-eGFP-PACT and mCherry-kendrin(2059-2398)-eGFP-PACT. Both localize to the centrosomes and enabled us to monitor local separase activity at the centrosome in real time. Both centrosomal sensors were cleaved by separase before anaphase onset, earlier than the corresponding H2B-mCherry-Scc1(142-467)-eGFP sensor at chromosomes. This indicates that substrate cleavage by separase is not synchronous in the cells. Depletion of the proteins astrin or Aki1, which have been described as inhibitors of centrosomal separase, did not led to a significant activation of separase at centrosomes, emphasizing the importance of direct separase activity measurements at the centrosomes. Inhibition of polo-like kinase Plk1, on the other hand, decreased the separase activity towards the Scc1 but not the kendrin reporter. Together these findings indicate that Plk1 regulates separase activity at the level of substrate affinity at centrosomes and may explain in part the role of Plk1 in centriole disengagement.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Paintrand M, Moudjou M, Delacroix H, Bornens M (1992) Centrosome organization and centriole architecture: their sensitivity to divalent cations. J Struct Biol 108: 107–128. - PubMed
-
- Tsou MF, Stearns T (2006) Mechanism limiting centrosome duplication to once per cell cycle. Nature 442: 947–951. - PubMed
-
- Fang G, Zhang D, Yin H, Zheng L, Bi X, et al. (2014) Centlein mediates an interaction between C-Nap1 and Cep68 to maintain centrosome cohesion. J Cell Sci 127: 1631–1639. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
