

Systematic analysis of Zn2Cys6 transcription factors required for development and pathogenicity by high-throughput gene knockout in the rice blast fungus
- PMID: 25299517
- PMCID: PMC4192604
- DOI: 10.1371/journal.ppat.1004432
Systematic analysis of Zn2Cys6 transcription factors required for development and pathogenicity by high-throughput gene knockout in the rice blast fungus
Abstract
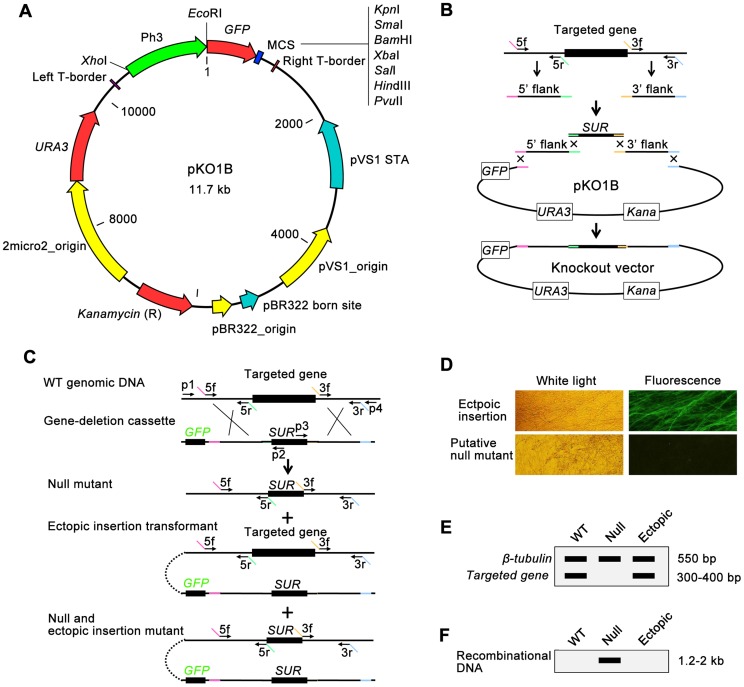
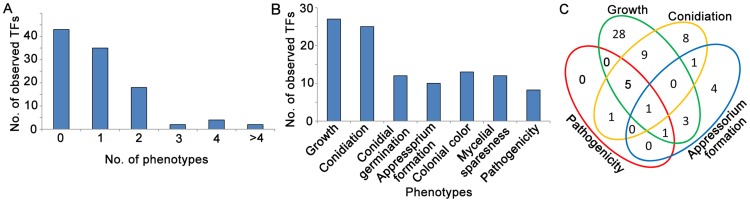
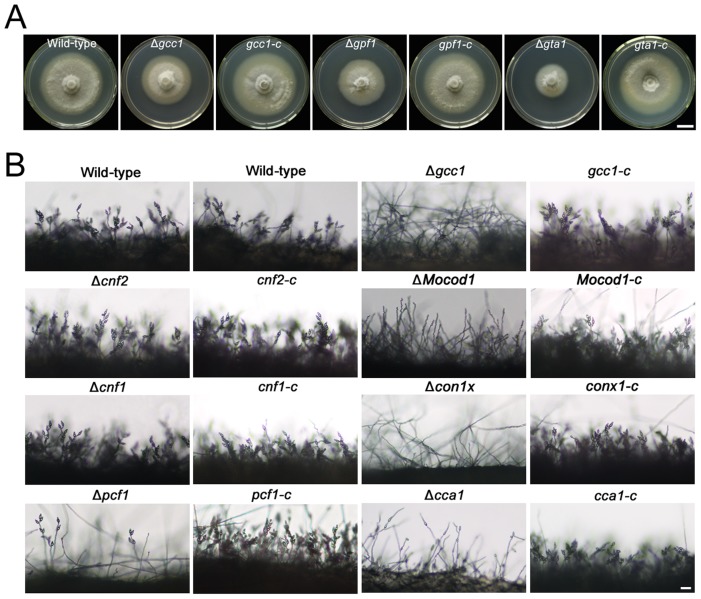
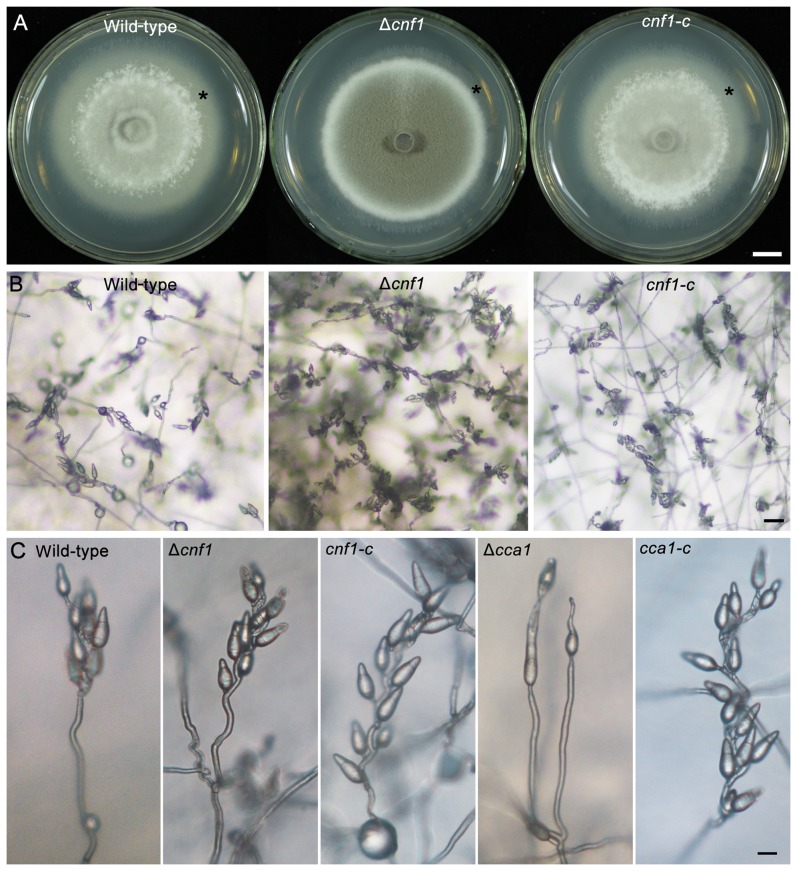
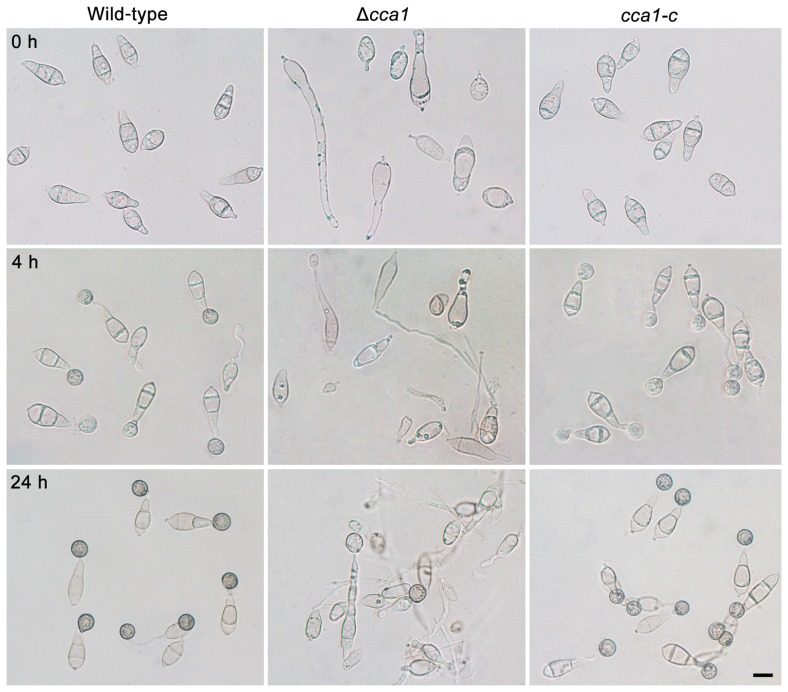
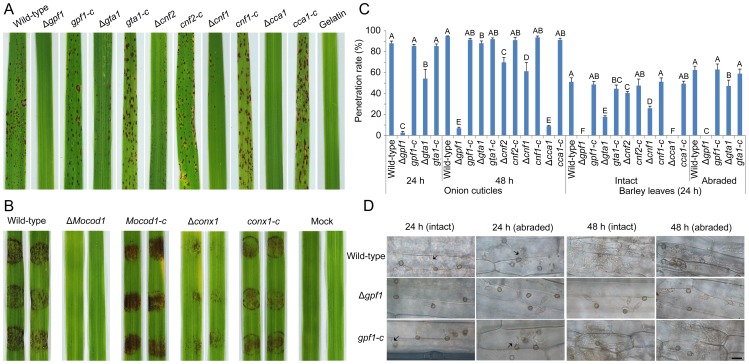
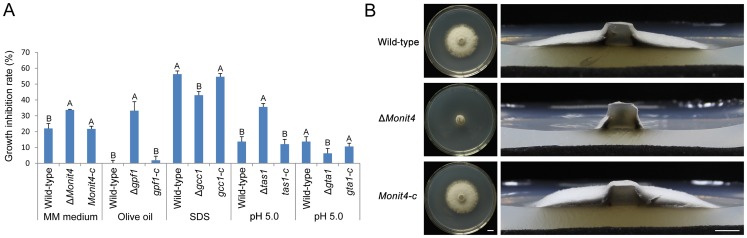
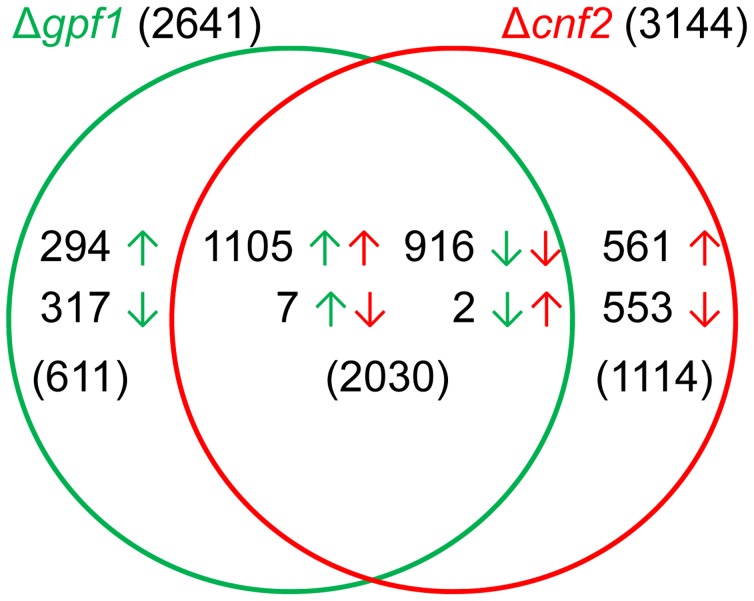
Because of great challenges and workload in deleting genes on a large scale, the functions of most genes in pathogenic fungi are still unclear. In this study, we developed a high-throughput gene knockout system using a novel yeast-Escherichia-Agrobacterium shuttle vector, pKO1B, in the rice blast fungus Magnaporthe oryzae. Using this method, we deleted 104 fungal-specific Zn(2)Cys(6) transcription factor (TF) genes in M. oryzae. We then analyzed the phenotypes of these mutants with regard to growth, asexual and infection-related development, pathogenesis, and 9 abiotic stresses. The resulting data provide new insights into how this rice pathogen of global significance regulates important traits in the infection cycle through Zn(2)Cys(6)TF genes. A large variation in biological functions of Zn(2)Cys(6)TF genes was observed under the conditions tested. Sixty-one of 104 Zn(2)Cys(6) TF genes were found to be required for fungal development. In-depth analysis of TF genes revealed that TF genes involved in pathogenicity frequently tend to function in multiple development stages, and disclosed many highly conserved but unidentified functional TF genes of importance in the fungal kingdom. We further found that the virulence-required TF genes GPF1 and CNF2 have similar regulation mechanisms in the gene expression involved in pathogenicity. These experimental validations clearly demonstrated the value of a high-throughput gene knockout system in understanding the biological functions of genes on a genome scale in fungi, and provided a solid foundation for elucidating the gene expression network that regulates the development and pathogenicity of M. oryzae.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ou SH (1980) Pathogen variability and host resistance in rice blast disease. Annu Rev Phytopathol 18: 167–187.
-
- Dean RA, Talbot NJ, Ebbole DJ, Farman ML, Mitchell TK, et al. (2005) The genome sequence of the rice blast fungus Magnaporthe grisea . Nature 434: 980–986. - PubMed
-
- Valent B, Chumley FG (1991) Molecular genetic analysis of the rice blast fungus, Magnaporthe grisea . Annu Rev Phytopathol 29: 443–467. - PubMed
-
- Talbot NJ (2003) On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea . Annu Rev Microbiol 57: 177–202. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
